The Aphids
SYSTEMATIC TREATMENT OF APHID GENERA
(in alphabetical order)
P
|
Pachypappa Koch |
Eriosomatinae: Pemphigini |
About 13 species mostly host-alternating between Populus and roots of Picea. The fundatrices lack wax glands and the alate spring migrants have forewings with once-branched media. Accounts are available from Heie (1980; Fennoscandia and Denmark), Danielsson (1976, gall generations in Sweden; and 1990a, root-feeding generations), Danielsson & Carter (1991; root-feeding generations in UK), Blackman et al. (2019b; the UK), G. Zhang & Qiao (1997c; China), G.Zhang et al. (1999c, China) and Smith (1974a; North America).
Pachypappa aigeiros Zhang Known from only one alata and two immatures collected on Populus cathayana in July in Shanxi, China (G. Zhang & Qiao 1997c). The gall is not described.
Pachypappa marsupialis Koch Forming large bag- or blister-like galls (fig. 131E), open to the underside, on leaves of Populus nigra in Europe (Austria, Estonia, Germany, Poland, Romania). The fundatrix is brownish green with white wax. Alatae leave the gall in June (Börner & Heinze 1957); they have a pinkish yellow abdomen and BL 3.1-3.5 mm (Szelegiewicz 1982). The secondary host is unknown. A subspecies, P. marsupialis lambersi, produces yellow blister-like galls on Populus spp. (laurifolia, suaveolens) in Japan and China (Aoki 1976, G. Zhang et al. 1985). It has dimorphic first instar larvae, one morph being migratory and leaving the gall to invade others (Aoki 1979). Alatae of ssp. lambersi migrate to an unknown secondary host in July. 2n=10 (for ssp. lambersi).
Pachypappa myrtilli Börner Apterae secreting wax; BL 1.2-1.4 mm. Described from roots of Vaccinium myrtillus in Germany.
Pachpappa pilosa (Zhang) Appearance in life is unknown, BL of emigrant alata c. 3.2 mm. On Populus sp. in Tibet (Zhang & Zhong 1981b; as Sigmacallis pilosa).
Pachypappa populi (Linnaeus) Forming large pale green, yellowish to slightly reddish blister-like galls (fig. 131F), open to the underside, on leaves of Populus tremula in Europe (Fennoscandia, Latvia, Poland, the Alps), Central Asia (Kazakhstan; Kadyrbekov 2017a), and also reported (on the closely related P. davidiana) from China (Zhang & Zhong 1985). The fundatrix is bluish grey-black, with sparse or no wax; BL 5.1-6.3 mm. Alatae, with olive green abdomen and BL c. 3.5 mm, migrate in early July in Europe, to found large colonies in flocculent wax on roots of Picea abies. Sexuparae appear in September, but anholocycly on Picea roots also occurs in southern Sweden (Danielsson 1990).
Pachypappa pseudobyrsa (Walsh) The fundatrix is wax-dusted, yellowish olive green with a blackish head; BL 4.0-4.7 mm. Its feeding causes a small blister-like swelling in the centre of the underside of the leaf near the mid-rib (see influentialpoints.com/Gallery). Its numerous progeny spread out in May-July along the main veins, developing into dusky-dark alatae, BL c. 3.5 mm. On Populus spp. (deltoides, fremontii, tremuloides) in eastern North America, where it is widely distributed, and also in Colorado (Palmer 1952, as Asiphum). The life cycle is unknown; the alatae presumably migrate to an unknown secondary host, although Smith (1974a) thought that it might be monoecious.
Pachypappa rosettei (Maxson) Feeding by the fundatrix (BL 2.5-2.8 mm, appearance in life unknown) causes bending of leaves and stunting of stems to form a rosette-like nest, as in the closely-related European species P. tremulae. On Populus tremuloides in Colorado and Utah, USA, and secondary host populations are now recorded from Washington and Alaska (Pike et al. 2012). Alatae (BL 2.2-2.5 mm) are produced in July. Host alternation to roots of Picea spp. has now been confirmed by DNA bar-coding, and the apterous exule (BL 0.8-1.4 mm) redescribed (Pike et al. 2012). Aphids on roots of Pseudotsuga menziesii have also been tentatively assigned to P. rosettei (Danielsson 1990a). Anholocyclic overwintering may occur on spruce roots (Pike et al. 2012). Aphids infesting the roots of potted conifers in Canada and identified as P. tremulae may also be this species.
Pachypappa sacculi (Gillette) The fundatrix is wax-dusted, slate grey with brown head; BL 4.5-5.0 mm. It forms a large bag-like gall on P. tremuloides, with edges and tip of leaf curled downward (Harper 1966; as Asiphum). In western USA (Arizona, Colorado, Utah, Washingtom, Alaska) and across Canada (British Columbia, Alberta, Quebec, Ontario and Nova Scotia). Progeny of the fundatrix are all alate, with dark olive-green abdomen, BL 3-4 mm. Host alternation to Picea spp. has now been confirmed by DNA bar-coding, and a redescription provided of the apterous exule (BL 0.9-1.8 mm) (Pike et al. 2012). Anholocyclic overwintering may occur on spruce roots (Pike et al. 2012). 2n=10.
Pachypappa shaposhnikovi Gabrid Forming large bag-like galls from leaves of Populus spp. (talassica, laurifolia) in Kyrgyzstan (Gabrid 1989). BL of fundatrix c.4.8 mm, of spring migrant alata c.5 mm). The secondary host is unknown.
Pachpappa tortuosae Zhang Only known from a presumably vagrant sexupara collected on Salix matsudana var. tortuosa in China (G. Zhang & Qiao 1997c), and perhaps belonging to a species already described from Populus.
Pachypappa tremulae (Linnaeus) The fundatrix on twigs of Populus tremula is very large, almost globular, dirty reddish or yellowish brown, without wax, but appearing silvery due to a covering of fine hairs; BL 5.0-6.6 mm. Its progeny move onto new shoots and cause bending of leaves and petioles and stunting of growth of shoot, forming a rosette-like leaf-nest (see influentialpoints.com/Gallery). Second generation are all alate, orange-yellow to pinkish brown, wax-covered, with black head and pterothorax; BL 3.2-4.4 mm. They migrate in June to found colonies on roots of Picea abies (Stroyan 1975). Apterous exules are pale yellowish white with tufts of wax posteriorly; BL 1.3-2.1 mm. Sexuparae return to aspen in September-October, but populations also overwinter anholocyclically on roots of Picea. Widely distributed in the northern palaearctic, east to China (Zhang & Zhong 1985b) and Japan (Aoki 1976; as P. shirobamba). The numerous records from North America on roots of Picea spp. and Pseudotsuga menziesii (also as Asiphum tremulae and Rhizomaria piceae; Lavallee 1987) may all be due to confusion with P. rosettei (Footit et al. 2006). The male genitalia were described and illustrated by Wieczorek et al. (2012). 2n=10.
Pachypappa vesicalis Koch Forming irregular green to yellowish bag-like or blister-like galls (see Carter & Danielsson 1993 for photograph) on leaves of Populus spp. (alba, canescens). The fundatrix is reddish-yellow to orange-red, wax-dusted; BL 4.2-4.9 mm. All second generation are alate, with reddish-brown abdomen, BL c. 3.3 mm, migrating in June to found colonies on roots of Picea spp. (abies, glauca, sitchensis), with a record also from Pseudotsuga menziesii in Czech Republic (Hochmut 1968). Apterous exules on spruce roots are pale yellowish white secreting white filamentous wax. Sexuparae return to Populus in September-October; mating and oviposition on poplar bark were described by Carter & Danielsson (1991). In northern and north-west Europe.
Pachypappa warshavensis (Nasonov) (= varsoviensis Mordvilko) Forming loose leaf-nest galls on Populus spp. (alba, canescens, euphratica, pruinosa; see Carter & Danielsson 1993 or Casiraghi et al. 2019 for photographs). Fundatrices (feeding on twigs) are red-brown; BL 3.5-4.4 mm. Alatae from galls are yellowish orange, BL 3.0-3.8 mm, migrating in June, and apparently found colonies on roots of Populus and Salix spp., although this still needs to be confirmed by experimental transfers. Casiraghi et al. (2019) provide a redescription and illustrations of fundatrices and alate fundatrigeniae. Apterae tentatively identified as this species have been found on the roots of Salix caprea (Szelegiewicz 1982) and S. silesiaca (BMNH collection) in Poland, and on Populus alba roots in Sweden (BMNH collection). Carter & Danielsson (1993) found many adult apterae and dark yellowish alatoid nymphs associated with wax particles in soil around roots of P. canescens at a location in Wales in October, and collected adult sexuparae from the rough bark of the same group of trees. They also reported finding apterae in June on fine roots of P. canescens in England (Lancashire). In Europe and Central Asia (Kazakhstan: Kadyrbekov 2011a). 2n=10*.
|
Pachypappella Baker |
Eriosomatinae: Pemphigini |
One or two species closely related to Pachypappa, but the fundatrix has wax glands on the thorax and abdomen.
Pachypappella aliquiphila Zhang Known only from the fundatrix, BL 2.2 mm, collected from an unidentified Populus sp. in Shanxi Province, China (G. Zhang & Qiao 1997c). The gall is not described. Possibly this is a small specimen of P. lactea.
Pachypappella lactea (Tullgren) The gall is formed on Populus tremula by folding down of the leaf lamina to form a cone, which becomes bright orange. Fundatrices are greyish brown with white wax; BL 4.6-5.2 mm. All second generation are alate, with dark greyish brown abdomen, BL 2.5-3.5 mm, migrating in June-July to colonise roots of Picea abies (Danielsson 1990). The root-feeding apterae live in nests of dense wax wool, 1.5-2.0 mm across, usually one aphid in each (Albrecht 2015). Anholocyclic overwintering on Picea roots, including P. sitchensis (Danielsson & Carter 1991), may be common. Northern palaearctic and boreo-alpine in distribution, from Britain to east Siberia (Pashtshenko 1988a).
|
Paczoskia Mordvilko |
Aphidinae: Macrosiphini |
About 10 rather large, shiny brown species and subspecies mostly in eastern Europe and western Asia, related to Macrosiphoniella, with both pre- and postsiphuncular sclerites well-developed, and mostly associated with Echinops. The Echinops feeders have a long wedge-shaped R IV+V. Holman (1981) provided a key and detailed morphological comparisons, also noting the existence of a single aptera of an undescribed species from an unknown host in Ethiopia (BMNH collection, leg. H. Scott).
Paczoskia brevipilosa Tashev Apterae are shining dark brown; BL 1.9-2.8 mm. On leaves of Echinops spp. in Bulgaria and Rumania (Holman 1981). Biology has not been studied; Holman suggested that its morphology might indicate a close association with ants and subterranean life at some stage in its life cycle.
Paczoskia colchica Holman Apterae are shiny brown, BL 2.6-3.8 mm. On undersides of leaves of Echinops galaticus (= ossicus)in southern Russia (Krasnodar). Monoecious holocyclic, with oviparae and apterous males in late October (Holman 1981).
Paczoskia jurineicola (Bozhko) Apterae are shining dark brown with black appendages; BL c.2.2 mm. On Jurinea stoechadifolia in Ukraine, forming dispersed colonies on undersides of leaves (Bozhko 1976b).
Paczoskia longipes (Tashev) Apterae are shining dark brown, BL 4.3-4.7 mm. On Echinops spp.in eastern Europe (Bulgaria, Poland, Rumania).
Paczoskia major Börner Apterae are shining dark brown, BL 2.4-4.3 mm. Described from Echinops sphaerocephalus in Germany, and since found in various east European countries, and in Turkey and Iran (Goodarzifar et al. 2010). In Turkey it was common on stems of E. ritro (Remaudière & Tuatay 1963). A single aptera (without host data) ascribable to this species was collected in 1926 at 3,000 m in Ethiopia (BMNH collection, leg. H. Scott). Sexuales occur in Europe in September-October, with both apterous and alate males (original description). A form on E. microcephalus and Echinops sp. in Bulgaria and Turkey is regarded as a subspecies, P. major ssp. bulgarica Holman.
Paczoskia meridionalis Holman Apterae are shining dark brown, BL 3.1-4.2 mm. On Echinops spp. in Iran, Israel, Lebanon and former Yugoslavia. Holman (1981) compared the morphology of different populations.
Paczoskia obtecta Börner Plate 27e Apterae are shiny bronze brown, with bright red eyes, black siphunculi and cauda, antennae black except at base of III, and legs yellowish with dark segmental apices (see influentialpoints.com/Gallery); BL 3.1-4.3 mm. On undersides of lower leaves, and later on upper parts of stems, of Centaurea spp. In Europe (France, Sweden, Denmark, Germany, Poland, Austria, former Yugoslavia, Hungary, Czech Republic), and in Israel. Sexual morphs are apparently unknown. 2n=12.
Paczoskia paczoskii Mordvilko Apterae are shining dark brown, BL 2.3-4.2 mm. On stems and undersides of leaves of Echinops spp. in Europe (France, Rumania, Ukraine). Monoecious holocyclic; oviparae were recorded in France in October. Nevsky (1929a) described a subspecies from E. karatavicus in Uzbekistan, P. paczoskii ssp. turanica, and this subspecies isalso recorded from Kazakhstan (Kadyrbekov & Aoitzhanova 2005, Kadyrbekov 2014e) and Greece (Tsitsipis et al. 2007). Holman (1981) compared various populations and described P. paczoskii ssp. ruthenica from E. ritro ssp. ruthenicus in Hungary and Crimea. Kadyrbekov (2010, 2014g) identified aphids on E. albicaulis in Kazakhstan and on E. ritro in Russia as ssp. ruthenica.
Paczoskia wagneri (Remaudière & Tuatay) Apterae are shining dark brown to almost black in life; BL 2.7-3.4 mm. Described from an unidentified Echinops sp. in Turkey (Remaudière & Tuatay 1963, as Macrosiphoniella subgenus Paczoskia). No other morphs are recorded.
|
Paducia Koch |
Aphidinae: Macrosiphini |
A genus of one or two little-known species. Its taxonomic position within Aphidinae is uncertain.
Paducia antennata (Patch) (fig. 106A) Only oviparae (collected October-November) and a single alate vivipara have been described (Patch 1913, Hottes & Frison 1931). Oviparae are dark greenish or chocolate-brown to black dorsally, greenish yellow ventrally; BL c. 2 mm. Alata are mainly brown, BL c. 1.4 mm. Presumably it lives without host alternation on the roots of Salix sp., although the evidence for this host association is entirely circumstantial. Recorded from Illinois, New York and Maine, USA.
Paducia aterrima Hille Ris Lambers Apterae are “probably shiny black”; BL 2.0-2.2 mm. Other morphs are unknown. On roots of Salix sp. in Greenland (Hille Ris Lambers 1952). Some differences from P. antennata were given by Hille Ris Lambers, but as different morphs were being compared the distinctness of the two species requires verification.
|
Panaphis Kirkaldy |
Calaphidinae: Panaphidini |
Large distinctive aphids with short antennae living on Juglans. The three described taxa are closely related and could perhaps be geographic variants of a single species. In the past the name Callaphis was widely used for this genus, but was suppressed following a plea by Quednau (1983), on the grounds of potential confusion with Calaphis Walsh. Quednau (2003) revised the genus.
Panaphis juglandis (Goetze) All viviparae are alate, large and stout-bodied, with dark head and thorax, yellow abdomen with dark brown transverse bands, and forewing veins fuscous-bordered (see influentialpoints.com/Gallery); BL 3.5-4.3 mm. On upper sides of leaves of Juglans regia, in rows along the veins, often ant-attended. In Europe, central Asia, Pakistan and north-west India, and introduced (in 1928) to western USA, where populations seemed to increase after the successful control of Chromaphis juglandicola . It appears that these two species cannot readily coexist, because colonies of P. juglandis on upper surfaces of walnut leaves are adversely affected by a rain of honeydew from colonies of Chromaphis on the undersides of the leaves above (Olson, 1974). Oviparae and alate males occur in September-October. The male genitalia were described and illustrated by Wieczorek et al. (2011). Masoodi et al. (1987) studied its population dynamics in Kashmir. Biological control has been attempted in California with a Praon species from Afghanistan (Mackauer & Sullivan 1982). 2n=22.
Panaphis nepalensis (Quednau) All viviparae are alate, colour in life unrecorded but probably like P. juglandis (see Quednau 1973). On Juglans regia in Nepal, Tibet and China (Yunnan; Qiao et al. 2005b). Sexuales are unrecorded. Zhang in Zhang & Zhong (1985c) described a subspecies, P. nepalensis yunlongensis, from Juglans mandshurica in Yunnan, China.
|
Paoliella Theobald |
Lizeriinae |
About 22 species on Burseraceae and Combretaceae in Africa (plus one in India), closely related to the South American genus Lizerius. Both apterous and alate viviparae are known for most species, but there are a few species which do not seem to produce apterae. Apterae and often alatae have conspicuous long dorsal processes (fig. 40). Quednau (2010) reviewed the world fauna and illustrated all available morphs.
Paoliella ayari (Eastop) Apterae are yellow, BL c. 0.9 mm, on undersides of leaves and young shoots of Commiphora spp. (pedunculata, angolensis). Recorded from Nigeria, Tanzania, South Africa and Zimbabwe. Life cycle unknown.
Paoliella browni Quednau The single known aptera, possibly a fundatrix, was dark brownish velvety green with white dorsal processes; BL c. 1.1 mm. Quednau (2010) has illustrated the alata and alatoid nymph of this species (the two alatae described in Quednau (1962) are P. echinata). On Combretum gueinzii (= C. molle) in South Africa. Van Harten (1979) described an aphid with greenish yellow alatoid nymphs living solitarily on undersides of very old leaves of Combretum sp. in Angola as a subspecies, P. browni capinganae. Alate males and apterous oviparae of this form (the latter dark velvety green, BL 1.0-1.1 mm) were collected in early June. An alata trapped in Burundi (Remaudière & Autrique 1985) may be this subspecies.
Paoliella capeneri Quednau Apterae are unknown and colour of alatae in life unobserved; BL of alata 1.0-1.4 mm. On Commiphora angolensis in South Africa (Quednau 2010).
Paoliella chiangae van Harten & Ilharco Only alatae are described; they have head, thorax and legs brown-black, abdomen greyish brown with dark tubercles, BL 1.2-1.6 mm (van Harten & Ilharco 1972). Living singly or in small colonies on undersides of young leaves of Combretum sp. (?psidioides) in Angola.
Paoliella commiphorae (Doncaster) All viviparae are alate, yellowish, thickly dusted with grey wax; BL c. 1.5 mm. On Commiphora eminii ssp. zimmermannii in Kenya and Malawi. Similar aphids collected on C. fischeri (= mossambicensis)in Tanzania were described as a subspecies, C. commiphorae persimilis (Eastop 1955). Alate males of this latter form have been trapped in January-March (BMNH collection).
Paoliella commiphoroides Quednau Apterae are unknown, colour of alata in life was not observed; BL of alata 1.2-1.5 mm. On Commiphora mildbraedii in Kenya (Quednau 2010). Oviparae and alate males were collected in May along with alate vivparae and alatoid nymphs.
Paoliella delottoi (Hille Ris Lambers) Apterae are brownish, BL 0.9-1.2 mm, feeding on undersides of shoot-tips of Commiphora africana in Eritrea and Nigeria. The life cycle is unknown. Aphids collected on C. dulcis in Namibia (BMNH collection, leg. H.D. Brown) are also regarded as this species.
Paoliella eastopi Hille Ris Lambers Colour in life is unknown; BL of aptera 0.9-1.4 mm. Found in large numbers on Commiphora eminii ssp. zimmermannii in Kenya (Hille Ris Lambers 1973d). Alatae were present in January, March and May.
Paoliella echinata Eastop Apterae are black with white dorsal processes; BL c. 0.8 mm. Immatures are yellow. On Commiphora pilosa in Tanzania and South Africa. The biology is unknown.
Paoliella (Lizerocallis) flava (Sousa-Silva & Ilharco) Apterae are yellowish, with dark apices to antennae and rostrum; BL 0.6-0.8 mm. Living dispersed on undersides of leaves of Tibouchina granulosa in Brazil (Sousa-Silva & Ilharco 2003). Quednau (2010) illustrated apterous and alate morphs.
Paoliella harteni Ilharco Only alatae are known, orange-reddish appearing rose-coloured due to fine pulverulence; BL 1.5-1.9 mm. On both sides of leaves of Terminalia spp. (brachystemma, sericea) in Angola (Ilharco 1971). Van Harten & Ilharco(1972) described the alate male and apterous ovipara, collected in July. Viggiani (1987) described a new aphelinid parasitoid from this aphid.
Paoliella hystrix Theobald Plate 7e The colour of the aptera in life is not recorded; BL 0.5-0.8 mm. Alatae and life cycle are unknown; only known from original collection on Commiphora savoiae (= edulis ssp. boiviniana?) in Somalia (Theobald 1928).
Paoliella (Lizerocallis) laxutuberculata Quednau Apterae are yellowish; BL c. 0.9-1.0 mm. On Combretum laxum in Belize, and also found on an unidentified host in Mexico. Apterae and alatae were described an illustrated by Quednau (2010). [This aphid was wrongly identified as Lizerius cermelii in Blackman & Eastop (2006, p. 1189).]
Paoliella (Kenyaphis) lepidosoma Quednau Apterae are orange-coloured in life: BL 1.0-1.2 mm. On ?Commiphora sp. in Kenya (Quednau 2010). An immature ovipara was collected in May.
Paoliella longirostris Quednau Only alatae are known, and their colour in life is unrecorded, BL c. 1.5 mm. Described from an alata trapped in Zimbabwe (Quednau 1974), and subsequently collected in Burundi on Combretum sp. (Remaudière & Autrique 1985).
P. monotuberculata van Harten & Ilharco Dense colonies of reddish-brown alatoid nymphs together with a small number of adult alatae (BL 1.2-1.5 mm) were found on the undersides of young leaves of Combretum sp. (?psidoides) in Angola (van Harten & Ilharco 1972). The alata was illustrated by Quednau (2010), and data are tabulated for comparison with P. browni capinganae by van Harten (1979).
Paoliella nachensis (Eastop) All viviparae are alate, with head and thorax dark, abdomen yellow with dark dorsal processes; BL 1.1-1.4 mm. Living singly on undersides of leaves Combretum spp. (incl. apiculatum, ?psidioides). Recorded from Angola, Burundi and Tanzania. The life cycle is unknown.
Paoliella namaachae Ilharco Only alatae are known, with pale yellow-orange abdomen and slightly darker head and thorax; BL 0.9-1.2 mm. Living solitarily on undersides of leaves of Combretum molle in Mozambique. Ilharco (1976) discusses differences from the closely-related P. nachensis. The life cycle is unknown.
Paoliella nirmalae (David) Apterae are bright brick red with numerous dull white dorsal processes (see aphids of Karnataka website); BL of aptera 1.1-1.4 mm. On Terminalia arjuna in Mysore, India (David 1969a). Ghosh & Quednau (1990) provided a redescription of aptera and alata, and discussed the differences setting this Indian species apart from all the African ones. The life cycle is unknown.
Paoliella papillata (Hall) All viviparae are alate, very dark brown; BL 1.4-1.5 mm. Immatures are pale reddish brown with white dorsal processes (Hall 1932). On Combretum spp. (incl. apiculatum, molle), widely distributed in Africa south of the Sahara. Doncaster (1954b) described the alate male and apterous oviparae from amongst the type material, collected in Zimbabwe in December.
Paoliella pteleopsidis Quednau Only alatae are described; colour in life is unknown, BL maximally c. 1.8 mm. On Pteleopsis anisoptera in Angola (Quednau 1974).
Paoliella terminaliae (Hall) All viviparae are alate, brick red to very dark but with tip of abdomen always pale; BL c. 1.5 mm (Hall 1932). On Terminalia sericea in Nigeria and southern Africa (Angola, Malawi, South Africa, Zimbabwe). The life cycle is unknown.
Paoliella ufuasi (Eastop) Only alatae are known, appearance in life unknown; BL 1.2-1.4 mm. Described from trapped alatae in Tanzania (Eastop 1955), but apterae collected on Commiphora pilosa in Kenya are also identified with this species (Quednau 2010). The life cycle is unknown, but an aptera illustrated by Quednau and collected in September in Kenya might be a fundatrix.
Paoliella wettsteini Quednau Apterae are pale yellow with white dorsal processes; BL 1.0-1.2 mm. Alatae are very pale with head and thorax only slightly dusky. On leaves of Combretum gueinzii (= C. molle) in South Africa (Quednau 1964). The life cycle is unknown.
|
Papulaphis Robinson |
Aphidinae: Macrosiphini |
One fern-feeding North American species, related to Macrosiphum but with unusual antennal sensoriation. Alatae often have a once-branched media.
Papulaphis sleesmani (Pepper) Plate 26c (Fig.43b) Apterae are lemon yellow to lemon green; BL 1.6-2.1 mm. On Adiantum pedatum and Cystopteris bulbifera in eastern USA.
|
Parachaitophorus Takahashi |
Drepanosiphinae (?) |
A genus for one oriental species on Spiraea with apterae having reduced eyes (triommatidia in fundatrices and oviparae), a much shorter antennal PT than in alatae, and very long dorsal hairs arising from dark tuberculate bases. The life cycle is unknown, but it is believed to migrate to an unknown secondary host (see below). Formerly given its own subfamily (Parachaitophorinae), Sugimoto (2011c) placed this genus in Drepanosiphinae because of characters of the oviparous morph, although heteroecy is hitherto unknown in that subfamily.
Parachaitophorus spiraeae (Takahashi) (= P. sikhotealinicus Pashtshenko, = P. yamashitai Sorin; synonymies by Sugimoto 2011c) Apterae in life are brown with dark brown head, antennae, legs, siphunculi, cauda and dark brown spots at bases of dorsal hairs; BL 1.2-1.7 mm. On Spiraea spp. in Japan, Korea and east Siberia, forming compact ant-attended colonies on stems and at bases of young offshoots, but Japanese populations disappeared in late May perhaps indicating migration to an unknown secondary host. Alatae (migrants?) produced in spring and autumn were compared by Sugimoto (2011c), who also described oviparae and alate males collected on S. cantoniensis in October-November. The male genitalia were described and illustrated by Wieczorek et al. (2011).
|
Paracletus van Heyden |
Eriosomatinae: Fordini |
About four species resembling Forda except for the longer outer margin of the hind coxae. Only P. cimiciformis is known to complete the holocycle with galls on Pistacia; sexuparae of P. donisthorpei have been recorded from P. terebinthus (Roberti 1939, Nieto Nafría et al. 2002a), but the gall-living generations of that species are either unknown or confused with those of P. cimiciformis. Molecular studies indicate a close relationship between Paracletus and Forda compared with other genera of Fordini (Ortiz-Rivas et al. 2009). Hille Ris Lambers (1954b) discriminated the species on the material then available.
Paracletus bykovi (Mordvilko) Apterae are yellowish white; BL 2.4-3.5 mm. On roots of many species of Poaceae (Calamagrostis, Festuca, Hordeum, Setaria, Poa); also Luzula (Juncaceae). In eastern Europe (Poland, Ukraine), Turkey, Transcaucasia, Kazakhstan, and Uzbekistan, where a population was described as P. bykovi ssp. uzbekistanicus Kan. Apparently mainly or entirely anholocyclic, although Mordvilko (1935) described a sexupara collected on trunk of Pistacia mutica in Georgia. Different from other Paracletus in several respects, and possibly deserving of separate generic or subgeneric status (as Hemitrama).
Paracletus cimiciformis von Heyden The galls on Pistacia (khinjuk, palaestina, terebinthus, vera) are flat folds of the edges of the leaves, retaining the original leaf colour. The fundatrix – globose, yellow, with 6-segmented antennae, BL c. 1.5 mm – has never been properly described. Second generation are all alate (BL 1.7-2.4 mm, fig. 81N) emerging from the gall in September-October to found colonies on roots of many species Poaceae (e.g. Agrostis, Brachypodium, Dactylis, Festuca, Hordeum, Oryza, Poa, Polypogon, Triticum). Apterae of the typical form on grass roots are shining waxy yellowish white to pale brown, with body dorso-ventrally flattened, especially at lateral margins, and 6-segmented antennae; BL 2.4-3.5 mm (Plate 2d). They are always attended by ants, especially Tetramorium spp. However transfer experiments indicated the existence of a second distinct grass root-feeding apterous morph of this species (Ortiz-Rivas et al. 2009), and this has been confirmed by comparison of DNA sequences (Mróz & Depa 2012). This morph has a dusky olive- to apple-green, highly domed body, BL 1.8-3.3 mm, and 5-segmented antennae, and was previously identified as Forda rotunda Theobald. It has been recorded from many genera of Poaceae (Brachypodium, Dactylis, Festuca, Poa, Polypogon), and is also ant-attended (Zwölfer 1958, as Forda skorkini). The factors influencing the production of these two morphs and their inter-relationships have not yet been elucidated (see Salazar et al. 2105, supporting information). However, work in Spain has now established that the two morphs have different relationships with the ant Tetramorium semilaeve; the round morph has a normal mutualistic relationship, whereas the flattened morph is an aggressive mimic with cuticular hydrocarbons that resemble those of the ant larvae, so that it is transported into ants’ nests where it sucks on ant larval haemolymph (Salazar et al. 2015).
Sexuparae return to Pistacia in April-June; these and the sexual morphs were fully described by Roberti (1939). P. cimiciformis is recorded from Pistacia in southern France, Italy, Iran, Israel, Malta (Mifsud et al. 2009) and Turkey. Anholocyclic populations on roots of grasses, recorded as either typical P. cimiciformis or as the morph previously identified as Forda rotunda, occur outside the range of the primary hosts throughout Europe, in north Africa and across Asia to Japan, Korea and China. Some regionally variant populations have been described as subspecies (panicumi Bozhko, on Panicum miliaceum in Ukraine, and zhanhuanus Zhang & Qiao (Zhang & Qiao, 1998d) on Triticum aestivum in China). For fuller information see Zwölfer (1958) and references therein. T. Zhang et al. (1987) studied the bionomics of this aphid in China. Álvarez et al. (2009) studied the histology of the gall. 2n=16.
Paracletus donisthorpei Theobald Apterae are pale; BL c.2.9-3.0 mm. Described from nests of the ant Tapinoma nigerrimum, and since found on roots of Triticum sp. and unidentified woodland grasses (Nieto Nafría et al. 2002a). In Italy, Spain, Greece and Algeria. The life cycle is unknown; sexuparae have been collected on Pistacia terebinthus (Roberti 1939), but the galls have not been identified and are possibly identical to those of P. cimiciformis (Ortiz-Rivas et al. 2009).
Paracletus subnudus Hille Ris Lambers Colour of apterae in life is unknown; BL of last instar imm. 2.6-3.0 mm. On Hordeum sativum (= vulgare) in Israel. Recorded also now from Turkey (Şenol et al. 2017), but this requires additional confirmation.
|
Paracolopha Hille Ris Lambers |
Eriosomatinae: Eriosomatini |
Two species in east Asia similar to Colopha, but the media of the forewing of alatae is unbranched as in Tetraneura, and the apterae on the secondary hosts (bamboos) always have siphunculi, which are displaced towards the midline. The life cycle of only one species is known. Akimoto (1985a) gave a full account.
Paracolopha morrisoni (Baker) (fig. 125c) The gall on Zelkova serrata is a club-shaped outgrowth from the upper surface of the leaf lamina between veins, with a narrow stem and a swollen, roughened apex (http://www.agr.hokudai.ac.jp/systent/akimoto/gall/P.morrisoni.jpg and influentialpoints.com/Gallery). Alatae (fig. 120B) emerge in June-July and colonise roots of bamboos (Sasa spp.). Sexuparae return to Zelkova in September-October. Akimoto & Narita (2002) studied the influence of alternating temperatures on the hatchability of overwintered eggs. Moritsu’s (1958) record from Celtis sinensis var. japonica (as Tetraneura nishiyae) requires confirmation. Galls on Zelkova occur in Japan and southern China (as Tetraneura zelkovisucta; Zhang 1980b). Apterae on roots of bamboo are globular, pale yellow with greyish legs and antennae, covered completely with white filamentous wax (Malumphy 2012); BL 1.5-1.8mm. Presumably anholocyclic populations have been introduced into USA, where it is recorded from the roots of various bamboos (Phyllostachys, Arundinaria) in Maryland, South Carolina and southern California (Hille Ris Lambers 1966a). It has been recorded on bamboo roots in Britain since at least 1998, and may be widely distributed in Europe (Malumphy 2012). This species has a complex synonymy; commonly used names in the past have been Colopha moriokaensis (Monzen) and Dryopeia nishiyae (Matsumura 1931, nec. 1917). 2n=10*.
{kind=link}
Paracolopha takahashii Akimoto Appearance of apterae in life is unknown; BL 1.25-1.5 mm. On basal part of Carex breviculmis ssp. royleana in Japan. Only known from the original collection (1956).
|
Paradoxaphis Sunde |
Aphidinae: Aphidini |
Two species in New Zealand, resembling Aphis but lacking lateral tubercles on ABD TERG 7, and often also lacking them on ABD TERG 1. Molecular studies (von Dohlen & Teulon 2003, Foottit et al. 2008) indicate a close relationship with indigenous New Zealand Aphis species. Teulon et al. (2013) summarised collection data and other the available information about both species.
Paradoxaphis aristoteliae Sunde Apterae are reddish brown with black head and dusky thorax; BL of aptera 1.5-2.0 mm. On leaves of Aristotelia serrata in New Zealand (Sunde 1987). Oviparae and eggs were obtained in January (Teulon et al. 2013). Podmore et al. (2019) reported COI sequence variation between populations. 2n=8*.
Paradoxaphis plagianthi Eastop Apterae are salmon-brown to brown with a green to dark brown longitudinal stripe; BL 1.3-2.0 mm. On leaves of Plagianthus regius in New Zealand (Eastop 2001). Kean & Stufkens (2005) reported that Hoheria angustifolia and the woody vine Parsonsia heterophylla may act as “spill-over” hosts. It seems to be anholocyclic, persisting on plump terminal buds for the short time in winter when Plagianthus is completely leafless (Kean 2002).
|
Paramyzus Börner |
Aphidinae: Macrosiphini |
Three palaearctic species, one on Rosaceae and two on Apiaceae, resembling Myzus but with apterae having secondary rhinaria on ANT III. Shaposhnikov & Stekolshtshikov (1989) reviewed the genus and provided a key to species.
Paramyzus heraclei Börner Plate 18e Apterae are white or yellow, shiny, pale except for tarsi (see influentialpoints.com/Gallery); BL 1.3-1.9 mm. On undersides of basal leaves of Heracleum spp., causing many small yellow spots and slight vaulting of the leaves. In Japan it is recorded from Angelica spp. (Miyazaki, 1971). Monoecious holocyclic on Heracleum in Germany, with alate males; a full account of the life cycle and sexual morphs was given by Müller (1977b). Widely distributed in Europe (not recorded from Scandinavia), Turkey, and in east Siberia and Japan. Japanese populations are possibly a different race or subspecies, P. heraclei ssp. similis Takahashi (see also Shaposhnikov & Stekolshtshikov, 1989).
Paramyzus longirostris Miyazaki Apterae are pale yellow, yellow or green, with antennae black towards tips; BL 1.1-1.4 mm. Alatae are undescribed. On Potentilla spp. in Japan and east Siberia (Kamchatka), and also found on Fragaria sp. in Japan (BMNH collection, leg. R.L.Blackman). 2n=14.
Paramyzus sii Shaposhnikov & Stekolshchikov Apterae are shiny greenish yellow with black eyes and tarsi, greenish siphunculi and whitish cauda; BL 1.6-2.0 mm. Alatae are undescribed. On Sium suave, at leaf bases and on inflorescences, in east Siberia (Vladivostok).
|
Paranipponaphis Takahashi |
Hormaphidinae: Nipponaphidini |
One species on Quercus in Japan, close to Nipponaphis but without siphuncular pores.
Paranipponaphis takaoensis Takahashi Apterae are nearly circular, with side of body perpendicular (“pill box-shaped”), blackish brown, BL 1.5-2.0 mm. On branches of an unidentified Quercus sp. in Japan (Takahashi 1959b). Other morphs and life cycle are unknown.
|
Paraphorodon Tseng & Tao |
Aphidinae: Macrosiphini |
One species in China with Phorodon-like digitiform process on the antennal tubercles, and dorsal cuticle densely papillated.
Paraphorodon omeishanaensis Tseng & Tao Apterae are pale green, with dusky antennae; BL c.1.3 mm. Alatae are undescribed. On undersides of leaves of an unidentified Rubus sp. in China. This species has been mistakenly synonymised with Phorodon cannabis.
|
Parathoracaphis Takahashi |
Hormaphidinae: Nipponaphidini |
Seven species in east and south-east Asia differing from Thoracaphis in the complete fusion of prosoma and abdominal plate, different cuticular ornamentation, spine-like marginal hairs and absence of siphunculi. Six species are recorded from Fagaceae, although one of these was originally described from Litsea; the host of the seventh is only tentatively recorded as Myrica. Only apterous viviparae are known. Some of the species may have unrecognised gall-forming generations on Distylium. The genus was reviewed and keyed by J. Chen et al. (2016a), and two of the Chinese species were reviewed by Qiao et al. (2018).
Parathoracaphis cheni (Takahashi) Apterae are black, oval, flattened, strongly sclerotised, without wax: BL c. 0.9 mm. Sedentary on undersides of leaves of ?Myrica rubra (host is queried in original description) in China (Chekiang). The life cycle is unknown.
Parathoracaphis elongata (Takahashi) Apterae are black, elongate oval, strongly sclerotised, with numerous wax filaments dorsally: BL c. 0.5 mm. On undersides of leaves of Quercus sp., and other unspecified Fagaceae, in Thailand (Takahashi 1941). The life cycle is unknown.
Parathoracaphis gooti (Takahashi) Apterae are black, broadly oval, flattened, sclerotised, with much white marginal wax: BL c. 1 mm. On undersides of leaves of Quercus sp. in Malaya (Takahashi 1950). The life cycle is unknown.
Parathoracaphis kayashimai (Takahashi) Apterae are presumably black, elongate oval, strongly sclerotised, perhaps without wax (not stated); BL c. 0.5 mm. On undersides of leaves of Quercus sp. in Malaya (Takahashi 1950). The life cycle is unknown.
Parathoracaphis manipurense (Pramanick, Samanta & Raychaudhuri) Apterae are broadly oval, aleyrodiform, brown to black; BL 0.75-0.9 mm. On Litsea sebifera (= glutinosa)in Manipur, India (Pramanick et al. 1983), and also recorded from Castanopsis sp. (A.K. Ghosh 1988). Biology is unknown. (An apterous morph collected on Amaranthus in another locality was described as the paratype fundatrix of this species, but no life cycle observations or other evidence linking these two morphs was reported, and Amaranthus is a most unlikely primary host for a hormaphidine.)
Parathoracaphis setigera (Takahashi) (fig. 91F) Apterae are black, elongate oval, strongly sclerotised, without wax; BL 0.5-0.7 mm. In loose clusters on undersides of leaves of Quercus spp. (glauca, myrsinaefolia) and Lithocarpus sp. in Japan, China (Sikong) and Taiwan (Tao 1966). There are also records from Palaquium sp. (Sapotaceae) and Styrax suberifolia (vagrants?) in Taiwan (Tao 1999). In Japan apterae occur on Quercus throughout the year (Takahashi 1958a).
Parathoracaphis spinapilosa Chen, Jiang & Qiao Apterae are black, with a fringe of white curved wax filaments, sparser posteriorly; BL 0.6-0.8 mm (J. Chen et al. 2016a). In large colonies on the undersides of leaves of an unidentified Quercus sp. (and also on another unidentified species of Fagaceae). In China (Fujian and Yunnan). The life cycle is unknown.
|
Parathoracaphisella Pramanick, Samanta & Raychaudhuri |
Hormaphidinae: Nipponaphidini |
One Indian species on Quercus, apparently close to Metathoracaphis. Only apterous viviparae are known.
Parathoracaphisella indica Pramanick, Samanta & Raychaudhuri Apterae are dark brown, with margin darker, oval: BL c. 1.1 mm. On Quercus sp. at Manipur, India (Pramanick et al. 1983). Other morphs and biology are unknown.
|
Pasekia Aizenberg |
Calaphidinae: Myzocallidini |
Pasekia cocciferina (Quednau & Barbagallo) Alatae are pale yellow, with head and thorax pale brownish laterally, abdomen with rather large, pale brown, paired dorsal patches, antennae and legs mainly pale, and siphunculi pale; BL 1.4-2.4 mm. Immatures are pale yellowish with brown dorsal markings. On undersides of leaves of Quercus calliprinos, Q. coccifera and Q. ilex in the Mediterranean region (Israel, Lebanon, Sicily, France, Andorra, Spain). Sexuales are unknown; it seems to be anholocyclic, at least in Sicily (Quednau & Barbagallo 1991). 2n=14*.
Pasekia komareki (Pasek) Alatae are yellow, with head and thorax variably marked with black (typically there are fine, black lines on the prothorax, roughly in the shape of a pair of rectangles), and dorsal abdomen with a series of large, paired quadrate spinal sclerites, which have dark margins and paler central areas, and similar-sized but paler and more uniformly-pigmented marginal sclerites. BL 1.1-2.2 mm. On Quercus spp. (e.g. aegilops, cerris, coccifera, ilex, ithaburensis, petraea, pyrenaica, suber) in southern Europe, Mediterranean area and Middle East (Bulgaria, Czech Republic, Greece, Israel, Iran, Italy, Moldova, Turkey, Ukraine, Canary Islands, Andorra, Spain). Monoecious holocyclic; sexuales occur in November in Iran (BMNH collection, leg. S.H. Hodjat). 2n=14*.
Pasekia mediterranea (Quednau & Remaudière) Apterae are pale yellow to creamy white, with dark markings; BL 1.2-1.6 mm. On Quercus pubescens and Q. ilex in southern France and Spain (Quednau & Remaudière 1994a), and on Q. pubescens and related species in Italy (Sicily, Patti & Barbagallo 1997; Tuscany, Barbagallo et al. 2011). Specimens recorded as P. komareki from Q. pubescens and Q. canariensis in Sicily (Barbagallo & Stroyan 1982) are also referable to this species. A record from Turkey (Şenol et al. 2017) requires additional confirmation. Sexuales occur along with alate viviparae from November to January, their appearance probably depending on the timing of leaf senescence (Barbagallo & Stroyan 1982, as P. komareki).
Pasekia persica (Quednau & Remaudière) Colour of alata in life is unknown; BL 1.6-2.3 mm. On Quercus persicae in Iran and Turkey (Quednau & Remaudière 1994a). Sexual morphs occur in October.
Pasekia taurica (Quednau & Remaudière) Colour of alata in life is unknown, pronotum with four longitudinal dark stripes and dorsal abdomen with four longitudinal rows of dark sclerites; BL 1.2-1.8 mm. On Quercus spp. (calliprinos, ?robur) in Turkey and Greece. Sexuales occur in October-November (Quednau & Remaudière 1994).
|
Patchia Baker |
Calaphidinae: Myzocallidini |
One North American species on Fagaceae, perhaps closely related to Lachnochaitophorus, but the apterae have separate head and prothorax.
Patchia virginiana Baker Apterae are broadly oval, rather uniformly velvety black, secreting wax; BL 1.8-2.3 mm. Alatae are brown with a large black dorsal abdominal patch and thickly-bordered forewing veins. On bark of Castanea spp. in USA (Florida, North Carolina, Pennsylvania, Virginia). Apparently monoecious holocyclic; oviparae were collected in September (BMNH collection, leg. J.O. Pepper).
|
Patchiella Tullgren |
Eriosomatinae: Pemphigini |
Two palaearctic species host-alternating between Tilia and Araceae.
Patchiella kolokasia (Qiao & Zhang) Described from roots of Araceae in China, this species has since been found on Tilia in Japan (Kurosu et al. 2022) and Korea (Lee et al. 2026). It can be distinguished from Patchiella reaumuri by the relatively shorter antenna and relatively longer R IV+V.
Patchiella reaumuri (Kaltenbach) (fig. 115B) This species forms large leaf-nest galls on Tilia spp. in spring by twisting and stunting the terminal growth and clumping the leaves (see influentialpoints.com/gallery). Fundatrices are plump-bodied, yellowish brown to greenish; BL c. 3.5 mm. All second generation are alate, with abdomen green to yellow-green, BL 2.1-2.7 mm, migrating to found colonies on roots and tubers of Arum spp. (Roberti 1939) and other Araceae (Arisaema, Colocasia). Apterae on the secondary host are whitish, with a powdery wax coating, and flocculent wax on the posterior abdomen; BL 1.9-2.5 mm (Stroyan 1979b). It is recorded from Tilia in England, Italy, Bulgaria, Hungary, Spain, Greece and Turkey. Anholocyclic populations occur on roots of Araceae elsewhere (e.g. Hawaii, Solomon Islands). Pashtshenko (1984a) described a subspecies, P. reaumuri orientalis, from Tilia amurensis in east Siberia; this is presumably the insect referred to as orientalis Mordvilko in Shaposhnikov (1955).
|
Paulianaphis Essig |
Greenideinae: Schoutedeniini |
One species on Madagascar related to Eonaphis and Schoutedenia, but with two pairs of dorsal processes on abdominal tergite 7, and a different host association. Remaudière (1988) gave a full account.
Paulianaphis madagascariensis Essig Apterae are broadly oval (almost globular), their colour in life is not recorded; BL 1.7-2.1 mm. In galls on leaves of Apocynum (according to label on type slide in BMNH collection, although host is given as “Apocynaceae” in Essig (1958); host plant and locality data were given in error under Eonaphis pauliani (see Hille Ris Lambers 1966b). Alatae have thickly-bordered forewing veins. Only known from Madagascar. The life cycle is unknown.
|
Pehuenchaphis Mier Durante, Nieto Nafría & Ortego |
Aphidinae: Aphidini |
Two species in South America on roots of Senecio, related to Aphis but with reduced siphunculi placed on abdominal tergite 6, reduced triommatidia, an unusual distribution of lateral tubercles, and distinctive biology. One species is known only from trapped alatae (P. americana, previously described in the genus Seneciobium).
Pehuenchaphis agilissima Mier Durante, Nieto Nafría & Ortego Apterae are dark green to brown, sometimes wax-dusted; BL 1.5-2.4 mm. Apterae have secondary rhinaria distributed III 14-18, IV 0-10. On Senecio spp., living in small ant-attended groups on thick, lignified roots, and at bases of lignified stems, moving quickly when disturbed. Monoecious holocyclic with apterous males (Mier Durante et al. 2003).
|
Peltaphis Frison & Ross |
Saltusaphidinae |
One species in North America distinguished by its dark sclerotic carapace, with abdominal tergites 2-6 fused together, and by the long dorsal hairs which often have furcate apices.
Peltaphis hottesi Frison & Ross Apterae are dark brown, almost black; BL c.2.1-2.3 mm. On Carex spp., only known from Illinois and Wisconsin, USA and Quebec, Canada. Alatae are unknown. Monoecious holocyclic, with oviparae and apterous males in October-November (original description).
|
Pemphigus Hartig |
Eriosomatinae: Pemphigini |
Over 70 species have been described, of which 45 are known to form galls on leaves or twigs of Populus (subgenus Eupopulus). Alatae emerging from these galls have a black head and pterothorax and a rather elongate, yellow-green to grey-green, wax-dusted, abdomen. The life cycles of 18 of these gall-formers are known; 4 are monoecious on Populus, the alate sexuparae being produced in the galls, and in the other 14 the alate progeny of the fundatrix migrate to various herbaceous mainly dicotyledonous secondary hosts in Amaranthaceae, Apiaceae, Brassicaceae, Compositae/Asteraceae, Euphorbiaceae, Leguminosae/Fabaceae and Ranunculaceae, and where they found colonies on the roots, or occasionally in woolly wax masses above soil level. Most of the species known only from their root-feeding generations feed on herbs in the above-named families, but at least one species feeds on Salix roots. Many species are recognised as distinct only from the morphology of the galls and emigrant alatae produced on Populus, or by experimental host transfers that show specificity to particular secondary hosts. Identification of apterae from roots using morphological criteria is generally difficult or impossible, so host records to species level based on apterae alone are almost always suspect. Zwölfer (1958) provided morphometric parameters for populations of unidentified Pemphigus spp. on roots of Poaceae and various dicots in southern Germany. Biology and ecology of most root-feeding populations are little-known; a Pemphigus species from Gnaphalium in India (western Himalaya; leg. S Chakrabarti), sent to BMNH for karyotyping in 1989 was heavily parasitised by an unidentified mermithid parasite (Blackman& Eastop 2006, p.1261; this is the first published record of a mermithid-aphid host association).
The species are distributed rather evenly throughout the northern hemisphere; 17 are described from Europe, 10 from the Middle East and Central Asia, 16 from east Asia, and 21 from North America. Accounts are available for Britain (Furk & Prior 1975); European gall-makers on Populus nigra (Lampel 1960); Fennoscandia and Denmark (Heie 1980b); the UK (Blackman et al. 2019b); the Iberian peninsula (Nieto Nafría et al. 2002a); India (A.K. Ghosh 1984b); East Siberia (Pashtshenko 1988a; Babichev & Kirichenko 2020); Japan (Aoki 1975); China (Zhang & Zhong 1979c, G. Zhang & Zhong 1983b, G. Zhang et al. 1999c); and North America (Palmer 1952, Harper 1959a, Lange 1965, Smith 1985). Dunn (1960a) and Faith (1979) investigated gall formation. Halaj & Osiadacz (2013) reviewed the European gall-forming species and provided an illustrated key to their galls on Populus. Alvarez et al. (2020) compared the histology of seven Iberian species and related it to their phylogeny. N. Pike et al. (2007) published a phylogeny for nine species based on mtDNA sequences. A recent study by L. Liu et al. (2014) of relationships within and between Pemphigus and allied genera using mitochondrial, nuclear and symbiont DNA has shown that the genus is paraphyletic, with P. matsumurai and P. sinobursarius forming a separate clade basal to one that includes five other Pemphigus plus Thecabius and Epipemphigus.
Pemphigus betae Doane (= balsamiferae Williams) Galls are yellowish green sometimes tinged with red, pocket-like, formed usually on the underside of the poplar leaf near its base, on or adjacent to the mid-rib, with little or no deformation of the leaf (Harper 1959a, Whitham 1978). The opening can be either on the upper side of the leaf or underneath along the mid-rib. Primary hosts are Populus angustifolia, P. balsamifera, or occasionally P. deltoides ssp. occidentalis or P. trichocarpa. Alatae (BL 1.5-2.1 mm) leave galls in late June-early August (Harper 1959b), to colonise roots of Beta vulgaris, and also perhaps on other Amaranthaceae (formerly Chenopodiaceae) such as Chenopodium album, and possibly Spinacia. Apterae on chenopods are yellowish aphids secreting white wax, with BL 1.9-2.4 mm. Anholocyclic populations can persist all year on secondary hosts (see Moran & Whitham 1988), and are widely distributed in North America. The life cycle was studied by Harper (1963) in southern Alberta, and by Moran & Whitham (1988) in Utah. This aphid’s evolutionary ecology has been studied in some detail, including the territorial behaviour of first instar fundatrices in selection of galling sites (Whitham 1979), population dynamics in relation to galling site (Whitham 1978, 1980), genetics of resistance in the host plant (Paige & Capman 1993), induction of sexuparae (Moran et al. 1993a), and discrimination between host trees by returning sexuparae in autumn (Moran & Whitham 1990). However, interpretation of all this work is problematic as Foottit et al. (2010a) have demonstrated that there is a complex of Pemphigus species in western North America sharing the same cottonwood primary hosts. Taxonomic affinities with palaearctic populations of Pemphigus on chenopod roots (see P. fuscicornis) are unclear. See also Blackman & Eastop (2000).
Pemphigus birimatus Ivanoskaya Galls on Populus laurifolia in northern Kazakhstan are irregular in shape, with two slit-like openings, and are formed on the leaf petiole (Ivanoskaya 1973). Also recorded on P. talassica, and from western Siberia (Kadyrbekov 2017a) and eastern Siberia (Babichev & Kirichenko 2020). The emigrant alata (BL 2.8 mm, with strap-like secondary rhinaria distributed III 11, IV 5, V 4, VI BASE 7), could not be included in the key on the basis of the published description.
Pemphigus borealis Tullgren Galls on Populus laurifolia are green, elliptical, hazelnut-sized, usually in groups of 2-10 on stem at bases of twigs or leaf petioles (fig. 131K). P. laurifolia seems to be the preferred host (Dolgova 1970), although there are also records from P. balsamifera (planted), P. iliensis, P. koreana, P. nigra, P. simonii and P. suaveolens. The fundatrix is grey-green with slight yellowish tinge. Alatae (BL 1.5-2.5 mm) emerge in June-August and migrate to found colonies on the roots of Bidens spp., these colonies consisting only of immature sexuparae (Dolgova 1970). Adult alate sexuparae are yellowish, with tufts of wax, BL c.2 mm, and migrate back to Populus in August-September. P. borealis occurs throughout the northern palaearctic, from Sweden to China and Japan. A record from England is possibly ascribable to P. trehernei (Foster 1975). The recorded occurrence of root-feeding populations in Idaho, USA (Griffiths et al. 1976) also requires confirmation. In east Asia there is probable confusion with P. dorocola (q.v.), which is described from very similar galls on P. maximowiczii (=P. suaveolens)and is possibly a synonym (Aoki 1975). P. yangcola, described from Populus sp. in China (Zhang & Zhong 1979c), is also very similar and a possible synonym. 2n=20.
Pemphigus brevicornis (Hart) Apterae are pale, with wax; BL c.1.8 mm. Described from apterae on corn (Zea) roots in Illinois, USA, and since recorded from roots of various plants in Illinois (Hottes & Frison 1931) and other states, but there are no good criteria for identification. Alate sexuparae have secondary rhinaria distributed III 5-7, IV 1-4 (according to Hottes & Frison 1931).
Pemphigus bursarius (Linnaeus) Poplar‑Lettuce Aphid, Lettuce Root Aphid Galls on Populus spp. (most commonly P. nigra) are yellowish or reddish when mature, purse-shaped, always formed on leaf petioles (fig. 131L, and see influentialpoints.com/Gallery). The fundatrix is greyish green, slightly wax-dusted. Alatae (BL 1.6-2.5 mm) leave the gall through a lateral opening over an extended period from late May to September, with peak emergence in late June-July (northern hemisphere). They found colonies mainly on roots of Compositae/Asteraceae (e.g. Cichorium, Lactuca, Lampsana, Sonchus, Taraxacum, Tussilago). Many of the records of P. bursarius from roots of plants in other families may be referable to other Pemphigus spp. Apterae on roots of secondary hosts are yellowish white with a tuft of white wax on the posterior part of the abdomen; BL 1.6-2.5 mm (Plate 2g). Alatae from root‑feeding colonies (sexuparae) have a brownish‑orange abdomen and secondary rhinaria distributed III 5-10, IV 1-4. Return migration of sexuparae to poplar is in October-September, but anholocyclic overwintering on roots of secondary hosts is common. P. bursarius occurs in Europe, across Asia to eastern Siberia (Babichev & Kirichenko 2020), northern and southern Africa, North and South America, and (perhaps) Australia and New Zealand. There is some uncertainty about its occurrence in Australia and New Zealand because, although bursarius-like aphids occur in bursarius-type galls on Populus and are also recorded from Sonchus roots, the secondary host populations are mainly recorded from Amaranthaceae and Polygonaceae. Miller et al.. (2005) found genetically distinct populations on different secondary host plants, indicating a complex of races and/or subspecies. Dunn (1960a) studied gall formation and (1960b) reviewed natural enemies. Alleyne & Morrison (1978a, b) gave accounts of its biology and natural enemies in Quebec. See also Blackman & Eastop (2000). 2n=20.
Pemphigus circellatus Zhang & Zhong Large galls are formed on the branches of Populus davidiana in China (Yunnan). Emigrant alatae (BL c. 1.8 mm) were collected in May (Zhang & Zhong 1985a). The life cycle is unknown.
Pemphigus cylindricus Zhang Only fundatrices are described, from a gall on an unidentified Populus sp. in Tibet (Zhang & Zhong 1981).
Pemphigus dorocola Matsumura The galls on twigs of Populus maximowiczii (=P. suaveolens) in Japan are pouch-shaped, reddish when mature, with a transverse apical slit (Aoki 1975). Also recorded from P. ciliata, P. koreana and P. suaveolens. The fundatrix is green. Alatae (BL 1.9-2.3 mm) emerge over an extended period (late July-November). The secondary host is unknown, and Chakrabarti (2001) suggested that it may not host-alternate. In Japan, Korea, east Siberia (Pashtshenko 1988a) and northern India (Chakrabarti 2001). Aoki (1978, 1980a) described aggressive and gall-cleaning behaviour by first instar larvae, and Aoki & Kurosu (1991d) described an aptera producing abnormal progeny in the gall. [P. dorocola is separated in the key to gall-formers on Populus according to the description by Aoki (1975), who does not recognise the presence of P. borealis in Japan. Alatae in the BMNH collection from galls on Populus suaveolens in Japan and Korea all have antennal sensoriation like that of P. borealis, and further work may show P. dorocola to be a synonym of P. borealis, as suggested by Takahashi (1920).] 2n=20.
Pemphigus echinochloaphaga (Zhang, Qiao & Chen) Appearance of apterae in life is unknown; BL 1.9-2.2 mm. On “grass” (Echinochloa sp.?) in China (Zhang et al. 1999, as Desiforda).
Pemphigus fatauae Shinji Apterae are undescribed, alatae (sexuparae?) from roots of Fatoua (as Fatauna) pilosa in Japan had secondary rhinaria distributed ANT III 6, IV 3-4, V 1. These may possibly have been sexuparae of Epipemphigus niisimae.
Pemphigus fuscicornis (Koch) Apterae are pale yellow or yellow-green to greyish green with wax tuft on posterior abdomen; BL 1.4-2.2 mm. On roots of Tripleurospermum inodorum (= Matricaria inodora) in Europe (UK, Denmark, Sweden, Germany, Finland) and Central Asia (Kazakhstan; Kadyrbekov 2017a). Originally also described from Chenopodium roots, and this name has therefore also been applied to very similar aphids found on Amaranthaceae (Atriplex, Beta, Chenopodium) in central and eastern Europe, and also in Iran and Central Asia (Kadyrbekov 2017a); however, these might be an anholocyclic population of P. betae introduced from North America (Blackman & Eastop 2000). Anholocyclic overwintering occurs on Tripleurospermum; alatae sexuparae (with secondary rhinaria distributed III 5-8, IV 1-3) are produced in autumn, but the sexual phase and primary host are unknown. Studies of microsatellite DNA variation in P. bursarius by Miller et al. (2005) may have included this species. 2n=20.
Pemphigus gairi Stroyan One of three palaearctic species producing similar pouch-shaped galls on or near the mid-rib on the upper sides of leaves of Populus nigra (incl. var. italica), with an opening to the underside of the leaf (the other two species are P. phenax and P. populinigrae). The fundatrix is dull green or greyish green under a thick covering of wax (see influentialpoints.com/Gallery). Alatae (BL 1.5-2.2 mm) emerge in June-July and migrate to colonise the roots of Aethusa cynapium (Stroyan 1964b). Apterae of the root-feeding generation are pinkish ochreous or pale straw-coloured; BL 1.3-2.0 mm. Sexuparae (with secondary rhinaria distributed III 4-7, IV 2-4) return in October. Described from England, and also recorded from Czech Republic, where Urban (2004) reported on the development and natural enemies of its gall generations on P. nigra, and from Ukraine. There seem to be no other literature records of this species, possibly due to confusion of its galls with those of P. phenax and P. populinigrae, as it is presumably more widely distributed in Europe.
Pemphigus hydrophilus Narzikulov Apterae are yellowish green, secreting flocculent wax; BL 1.9-2.2 mm. One alata (sexupara?) had secondary rhinaria distributed III 6, IV 5, V 2. On the moss Cratoneurum filicinum, and possibly also on Hygrohypnum luridum. In Tajikistan at 1600-1700 m.
Pemphigus ignotus Habib & Ghani Recorded as forming leaf galls on Populus ciliata in Pakistan (Habib & Ghani 1970), but no description is given, and this is therefore a nomen nudum.
Pemphigus immunis Buckton (= lichtensteini Tullgren) The galls are quite large (up to 4 cm maximum diameter), greenish brown or reddish, walnut-like in shape and surface texture, thick-walled, with an apical opening, on twigs or branches of Populus spp., especially nigra and euphratica (fig. 131M, and see influentialpoints.com/Gallery). The fundatrix is glaucous green (Roberti 1938). Alatae (BL 2.2-2.5 mm) emerge from late April to early August and found colonies of apterae (BL 1.2-1.8 mm) secreting copious wax on roots of annual Euphorbia spp. (helioscopia, peplus; Hille Ris Lambers 1973b). A.K. Ghosh (1984) reviewed its biology. In Europe, north Africa, south-west and central Asia, Pakistan, north-west India, and China (Tseng & Tao 1936; as P. napaeus). A record from Euphorbia in USA (as brevicornis) may also be this species. Habib & Ghani (1970) studied natural enemies in Pakistan, and Bhagat (1982a) reported a parasitoid and hyperparasitoid in Kashmir. 2n=20 (or 2n=10 according to Pal & Khuda-Bukhsh, 1982).
Pemphigus iskanderkuli Narzikulov Galls on Populus dentata when mature are pinkish to red, rounded, smooth, walnut-sized, with apical opening, situated on upper leaf surface at junction of petiole and lamina (Narzikulov 1957). BL of alata 2.7-3.5 mm. In central Asia (Tajikistan, Kirgizia). The life cycle is unknown. The alatae seem indistinguishable from those of P. napaeus, and Doncaster (1969) suggested that the two might be conspecific, but for reported differences in the morphology of the fundatrices, and position and colour of the galls. Populations in Kirgizia on P. laurifolia described by Gabrid (1989) as Pemphigus sp. aff. iskanderkuli would appear to be large specimens of iskanderkuli, although the galls on P. laurifolia illustrated by Gabrid appear very irregular in shape.
Pemphigus knowltoni Stroyan Galls are green, pouch-like, of variable shape, on undersides of leaves of Populus angustifolia alongside the mid-rib at the base of the lamina (Stroyan 1970). Fundatrix and immatures in gall are blackish green. Alatae (BL 1.8-2.1 mm) are mature in the gall in July, and migrate to an unknown secondary host. Described from Utah, USA.
Pemphigus laurifolia Dolgova Galls occur on Populus laurifolia in the Altai region of Russia. Alatae (BL c. 2.1 mm) emerge in July (Dolgova 1973). It has also been reported to occur on P. talassica in northern Tien-Shan (Kadyrbekov 2013f) and in eastern Siberia (Babichev & Kirichenko 2020). The original description is not available; this species is keyed on the basis of alate specimens (paratypes?) collected by Dologova on 1.vii.71 (BMNH collection, leg. D. Hille Ris Lambers). [Note that Pemphigus laurifoliae Holman & Szelegiewicz 1972 (nomen nudum) is P. mongolicus Holman & Szelegiewica 1974a)].
Pemphigus longicornus Maxson Galls are elongate, thin-walled, on upper sides of leaves of Populus deltoides. The fundatrix is pale yellowish white. Alatae (BL 1.7-2.1 mm) emerge in May-June and migrate to an unknown secondary host (Maxson 1923). In eastern and southern USA. [According to Smith (1985), there is also an undescribed Pemphigus sp. on P. deltoides in North Carolina that causes elongate to slightly globular galls on the upper sides of the leaves, and has alatae with antennae intermediate in length and antennal sensoriation between longicornus and populivenae.]
Pemphigus mangkamensis Zhang Described from apterous viviparae from an unidentified Populus sp. in Tibet (Zhang & Zhong 1981b), and said to be similar to P. chomoensis. If the specimens are not fundatrices this would indicate absence of host alternation.
Pemphigus matsumurai Monzen The gall is of globular or irregular shape, at base of mid-rib on upper side of leaf, concolorous with mid-rib or slightly reddish (Aoki 1975, Zhang & Zhong 1983b, Chakrabarti 2021). Recorded from Populus ciliata, P. koreana, P. laurifolia, P. nigra, P. talassica and P. suavolens. The fundatrix is green. Alatae (BL 1.7-2.7 mm, fig. 86C) migrate in June-July to colonise roots of Thalictrum spp. In east and Central Asia (Kazakhstan, northern India, Pakistan, Bhutan, Mongolia, east Siberia, China, Japan). Also in Tajikistan, as Pemphigus montanus, described from Populus tadzhikistanica (Narzikulov 1957), is clearly a synonym. Aoki & Kurosu (1989) studied the influence of parasitisation of the fundatrix on the development of the gall and subsequent generations, and Takada et al. (2010) studied the effects of a parasitoid and three hyperparasitoids. Bhagat’s (1982a) record of a parasitoid of P. immunis possibly should be referred to matsumurai, as this is the only species known to form leaf galls on P. ciliata. DNA studies by L. Liu et al. (2014) place this species, together with P. sinobursarius,in a basal position in a phylogeny that includes Thecabius and Epipemphigus. 2n=12 (an unusual chromosome number for a Pemphigus, confirmed for Japanese samples from Thalictrum).
Pemphigus microsetosus Aoki A large closed pouch-gall is formed as an outgrowth of the leaf of Populus suaveolens, taking in the whole leaf when mature; lignified galls remain on the tree after leaf-fall (Aoki 1975). The fundatrix is yellowish green. Alatae (BL 2.1-2.5 mm) are adult from late July and leave gall through several secondary exit holes, migrating to an unknown secondary host. In Japan, and since recorded from west Siberia (Stekolshchikov & Novgorodova 2015) and east Siberia (Pashtshenko 1988a, Babichev & Kirichenko 2020). The latter authors record the host in eastern Siberia as P. laurifolia. 2n=22 (Blackman 1986).
Pemphigus mongolicus Holman & Szelegiewicz The gall is globular, pouch-like, on upper side of leaf of Populus laurifolia near the base of the mid-rib; green, becoming tinged with red when mature (Holman & Szelegiewicz 1974a). The fundatrix is palish green, producing a generation of dark green apterae in the galls. Monoecious; alate sexuparae (BL 1.6-2.0 mm) leaving galls in early August give birth to sexuales in bark crevices on the trunk. In Mongolia and probably in eastern Siberia (Babichev & Kirichenko 2020). [“P. semenovi“, noted by Mordvilko (1935) as monoecious on P. suaveolens in Siberia but apparently a nomen nudum, is probably this species.]
Pemphigus monophagus Maxson The gall is globular, pale greenish yellow, consisting of a swelling of upper side of base of leaf lamina with petiole broadened and somewhat twisted, and a slit-shaped opening underneath between petiole and leaf (fig. 131N, and Maxson 1934). Fundatrix honey-yellow to pale olive-green, producing a generation of creamy white apterae in the gall. Monoecious, alatae (BL 1.4-1.9 mm) emerging from galls in August-September to deposit sexuales on the bark of the trunk. On Populus angustifolia, P. balsamifera and P. trichocarpa in western North America. Aoki & Kurosu (1988) discussed the evolution of monoecy in this species.
Pemphigus mordvilkoi Cholodkovsky Galls, formed on the twigs of Populus ciliata, are smooth, globular, sessile, yellowish green to dark greenish brown (Chakrabarti et al. 1985; Chakrabarti 2021). The fundatrix is pale green. Chakrabarti (2001) noted that its first instar progeny were dimorphic, one form apparently exhibiting the characteristics of “soldiers”. The life cycle is unusual, and requires further study. Fundatrices have a long life (Chakrabarti & Das 2014). In Pakistan, alatae emerge from the galls over an extended period from August until January of the next year (BMNH collection). According to A.K. Ghosh et al. (1981), the alatae emerging from galls just before onset of winter in north-west India hibernate in bark crevices of the same tree, or in other sheltered places; however, these alatae are not sexuparae, so the monoecious life cycle suggested by them seems unlikely. Apterous exules and sexuparae likely to be those of P. mordvilkoi have been collected in Punjab in April on roots of Helipterum (= Acroclinium) roseum (Compositae; BMNH collection, leg. A.S. Sohi); similar sexuparae have also been collected on other Compositae in other parts of northern India and Nepal in March-April (BMNH collection), and sexuparae and sexuales were observed on P. ciliata in Pakistan in April-May (Habib & Ghani 1970). The oviparae laid eggs which hatched in 6-9 days, and the fundatrices formed galls in May-June. Thus a two-year life cycle with alternation between P. ciliata and Acroclinum is indicated. 2n=20.
Pemphigus nainitalensis Cholodkovsky Galls are about the size and shape of a hazelnut, sessile, on twigs of Populus ciliata, which are thickened in the region of the gall (Cholodkovsky 1912). Only known from the original descriptions of the fundatrix and the alata, from galls collected in Uttar Pradesh, India.
Pemphigus napaeus Buckton Galls on twigs of an unidentified Populus sp. were smooth, roundish or of irregular shape, shining green “slightly veined with brown, and variegated with yellowish patches” (Buckton 1896); but see comments by Doncaster 1969, p. 162). In the Northern Areas of Pakistan (Gilgit). There are no other authenticated records, and contrary to some authors (eg. A.K. Ghosh et al. 1981), the host species is unidentified. Reports of napaeus in Taiwan (e.g. Tseng & Tao 1936) should probably be referred to immunis. Doncaster (1969) remounted and redescribed the type material. See also under P. iskanderkuli.
Pemphigus nortonii Maxson Galls on Populus deltoides especially var. occidentalis (= monilifera) are pale yellowish green, formed by a swelling at the base of the leaf lamina, twisted on the flattened, broadened petiole; resembling those of P. populicaulis (Palmer 1952), but with a long slit-like opening (Harper 1959a). Alatae emerge over an extended period (June-Sept; Harper 1959b, Grigorick & Lange 1968), migrating to an unknown secondary host. Apparently widely distributed in USA, and in Alberta, Canada. Apart from the greater number of rhinaria on ANT V and VI this species closely resembles P. junctisensoriatus. Populations from Alberta (Harper 1959a, b) seem to be intermediate in this respect, and the distinction between these two species needs to be verified.
Pemphigus obesinymphae Aoki & Moran Galls are formed on the petioles of late flushing leaves of Populus fremontii (Aoki & Moran 1994), and P. deltoides (Abbot 2009), usually close to the leaf blade. When mature they are almost spherical, 8-16 mm in diameter, with the lips of the transverse slit sometimes bulging outwards, and a circular ostiole about 1.2 mm in diameter somewhere along the slit. First instar progeny of the fundatrix in the gall are heavily sclerotised with well-developed fore legs and attack predators (Moran 1993). Abbott (2009) demonstrated using microsatellite DNA analysis that there was a high level of clonal admixture within galls, and Grogan et al. (2010) investigated its consequences. Alatae (BL 1.9-2.5 mm) fly in August-October, presumably migrating like P. populitransversus to cruciferous hosts (N. Chen et al. 2009), on which overwintering occurs. Sexuparae return to Populus in May of the following year, and the eggs hatch only a short time after oviposition. Apparently widely distributed in USA (Ohio, Tennessee, Arizona, Utah, Texas), and probably in northern Mexico. N. Chen et al. (2009) recorded molecular and phenological differences between P. obesinymphae and P. populitransversus on cabbage in Texas. This species was confused for many years with P. populitransversus (q.v.), and its distribution and range of both primary and secondary hosts requires further confirmation.
Pemphigus passeki Börner Galls are formed on leaves of Populus nigra (incl. var. italica) by swelling of the basal half of the mid-rib, the swelling being broadest towards the junction with the petiole (fig. 131O, and Prinsen 1990). Alatae (BL c. 2.1 mm) fly in June-August to colonise roots of caraway (Carum carvi). It is very similar to P. phenax but can be separated on the basis of morphological differences in the alate sexupara. In northern palaearctic; north-west, north and central Europe, Russian Altai region (BMNH collection, leg. L.P. Dolgova) and eastern Siberia (Babichev & Kiirichenko 2020). The spring migration was studied by Prinsen (1991). 2n=22.
Pemphigus phenax Börner & Blunck One of a complex of species in Europe producing similar mid-rib galls on upper sides of leaves of Populus nigra (incl. var. italica), with a slit-like opening underneath the leaf. The galls of P. phenax are reddish, tend to be larger than those of P. gairi, and more elongate (spindle-shaped) than those of P. populinigrae, somewhat more wrinkled, and often tinged with yellow laterally (Stroyan 1964b). The fundatrix is dull pale green when adult. Alatae (BL 1.5-2.4 mm) fly in June-August to found colonies on roots of wild and cultivated carrot (Daucus carota). Apterae on carrot are pale lemon yellow to yellowish‑white with white wax; BL 1.7-2.6 mm. Populations may also persist parthenogenetically on carrots remaining in the ground through the winter. In northern Europe (England, Denmark, France, Germany, Netherlands, Sweden), Turkey (Şenol et al. 2014) and west Siberia. For a more detailed account see Stroyan (1964b).
Pemphigus plicatus Dolgova Galls occur on Populus laurifolia in the Russia (Altai region, western and eastern Siberia, and are also recorded from P. laurifolia and P. nigra in Kazakhstan (Kadyrbekov & Aoitzhanova 2005). Alatae (BL c. 1.8 mm) were collected in July. The original description is not available. [It is included in the key on the basis of specimens collected by Dolgova on 1.vii.71 (BMNH collection, leg. L.P. Dolgova.)]
Pemphigus (Pemphiginus) populi Courchet Galls on Populus spp., especially P. nigra (incl. var. italica) are globular outgrowths of the mid-rib, not larger than a hazelnut, becoming club-shaped with the basal part narrower than the apex when mature, situated near the base on the upper side of the leaf (see influentialpoints.com/Gallery). Alatae (BL 1.4-2.5 mm, fig. 86D) emerge in June-July through a secondary opening at the top of the gall, migrating to found colonies on roots of Melilotus altissima (Furk & Prior 1975), and certain other Leguminosae (Lathyrus, Medicago). Apterae on secondary hosts are pale yellow-green, with white wax; BL 1.9-2.6 mm. Sexuparae (with secondary rhinaria distributed III 4-7, IV 1-2) return to P. nigra in September-October. In Europe, south-west and central Asia, and eastward to east Siberia (Babichev & Kirichenko 2020) and China (Zhang & Zhong 1979c). Furk & Prior (1975) reported parasitisation of fundatrices by Monoctonia pistaciaecola. Coffin & Remaudière (2001) compared the life cycle and morphology with P. vesicarius. N. Pike et al. (2007) studied the phenology of the gall generations. For a general account see Osiadacz & Halaj (2014).
Pemphigus populicaulis Fitch The gall is yellowish green to creamy, formed by swelling of base of leaf lamina, twisted around broadened petiole, rather like that of P. nortonii and P. populiglobuli but with a small, rounded exit hole on the underside (Palmer 1952, Harper 1959a). Especially associated with P. deltoides and P. deltoides ssp. occidentalis (= monilifera), but also recorded from other cottonwoods and P. tremuloides. The fundatrix is pale yellowish to whitish green.  (see influentialpoints.com/Gallery); Alatae (BL 1.7-2.0 mm) emerge in July-September. The secondary host has not definitely been established; Glendenning (1924) reported that it was Oenanthe sarmentosa, but did not provide evidence for this assertion. Hottes & Frison (1931) suggested that P. brevicornis Hart, described from corn roots, was the sexupara of populicaulis. Widely distributed in North America. Setzer (1985) studied gall mortality. Sokal et al. (1980) and Bird et al. (1982) studied geographical variation in eastern North America using multivariate techniques.
Pemphigus populiglobuli Fitch The gall is green, often tinged with red and/or yellow when mature; globular, conical or bullet-shaped, formed by swelling of base of leaf lamina and thickening of petiole, the exit hole being a slit on the underside between leaf and petiole (Maxson & Knowlton 1929). Primary hosts are Populus angustifolia and P. balsamifera, more rarely P. trichocarpa. The fundatrix is variously reported to be dark green (Palmer 1952) or pale yellowish white (Maxson & Knowlton 1934). Alatae (BL 1.5-2.3 mm) leave gall in June-early July (Alberta; Harper 1959b) for an unknown secondary host. Widely distributed in North America. Differences from P. populicaulis and other closely-related species are discussed by Maxson & Knowlton (1929).
On roots of unidentified Poaceae in Greenland, Iceland, and Svalbard, where it is represented by an anholocyclic secondary generation Wieczorek et al. (2024) formerly known as P. groenlandicus (Rübsaamen). Zwölfer (1958) noted that populations on roots of various grass species in Germany (Agrostis, Arrhenatherum, Avena, Cynosurus) agreed well with this species. There are subsequent records from France and Spain (Nieto Nafría et al. 2002a), the morphology agreeing best with that of the Greenland population (described as P. groenlandicus crassicornis Hille Ris Lambers). Differences from P. similis Börner, recorded from ?Agrostis alpina, Nardus stricta and Poa laxa in the Austrian Tyrol, and from P. vulgaris Raychaudhuri, Pal & M.R. Ghosh, described from Triticum aestivum and unidentified grasses in Sikkim, and West Bengal, India, have not been established.
Pemphigus populinigrae (Schrank) (= filaginis Boyer de Fonscolombe) Galls on Populus spp., especially P. nigra, are pouch-shaped (fig. 131P), similarly situated to those of P. phenax and P. gairi, on upperside of leaf lamina alongside mid-rib with opening along original line of invagination on underside of leaf. Gall formation was described by Dunn (1960). Mature galls are rounded and smooth on top, and of a dull reddish colour without much tinge of yellow (Stroyan 1964). The fundatrix is dark green. Alatae (BL 1.8-2.9 mm) emerge in June-August and migrate to found colonies above ground on Filago, Logfia and Gnaphalium, often in the tomentum of the inflorescences. Apterae on secondary hosts are yellow-green, with white wax-wool; BL 2.0-2.2 mm. Sexuparae (with secondary rhinaria distributed III 7-9, IV 2-4) return to P. nigra in September-October, but anholocyclic overwintering apparently occurs where the primary host is not available (Mordvilko 1935). Throughout Europe and in north Africa, south-west Asia, Central Asia and across to east Siberia (Pashtshenko 1988a). Records from China (Tao 1958) may be due to confusion with Epipemphigus niisimae. 2n=22.
Pemphigus populiramulorum Riley Galls are formed by swelling of new growth of twigs, or rarely at the bases of petioles; they are quite large (up to 2.5 cm diam.), rounded, green to brownish, with a transverse or vertical slit-shaped opening which often has protruding lips. The primary host is usually Populus deltoides ssp. occidentalis (= monilifera), but it is also recorded from P. balsamifera and P. fremontii. The fundatrix is pale creamy yellow (Palmer 1952). Alatae (BL 2.2-2.5 mm) leave galls in July-September (Harper 1959b), flying to an unknown secondary host. In western North America (Utah, Colorado, South Dakota).
Pemphigus populitransversus Riley Plate 6c The gall is formed by swelling and bending of petioles of the leaves of Populus deltoides (incl. ssp. occidentalis) and P. fremontii; it is green, globular to ovoid, with a usually transverse slit (see influentialpoints.com/Gallery). Sometimes recorded from other Populus spp. (acuminata, angustifolia, balsamifera, nigra, tremuloides, trichocarpa) However, some of these records, particularly those from western cottonwoods, may apply to other species. The fundatrix is whitish to pale dirty greenish yellow (Palmer 1952). Alatae (BL 1.7-2.0 mm) emerge in July and found colonies of pale yellow aphids in flocculent wax on roots of Brassicaceae (Jones & Gillette 1918), where it can be a serious pest (Royer & Edelson 1991). Sexuparae return to cottonwood in September-October to produce sexuales which lay overwintering eggs on the bark. Geographic variation in eastern USA has been studied extensively using multivariate methods (Sokal & Riska 1981). Bingham & Sokal (1986) studied host tree effects on morphology. P. populitransversus is recorded throughout most of North America east of the Rocky Mountains. However it is still unclear whether the name is being applied to a single species; apart from the confusion with P. obesinymphae (q.v.), Bird et al. (1979) found that two forms with different gall shape and phenology, and corresponding morphological differences in fundatrices and emigrant alatae, occur sympatrically on P. deltoides over a large area of the USA east of the continental divide. It should be noted that the morphology of their aphids from globular galls on late flushing leaves does not correspond with that described for P. obesinymphae.
Pemphigus identified as P. populitransversus have been found on Populus sp. in Mexico (BMNH collection, leg. R. Peña) and Chile (BMNH collection, leg. G. Monsalve); it has also been introduced to South Africa (Müller & Scholl 1958), where it occurs on Populus nigra (BMNH collection, leg. E McC. Callan) and P. deltoides, and the Azores (on Populus sp.; BMNH collection, leg. A. van Harten). Presumed anholocyclic populations also occur in New Zealand on Eutrema wasabi roots (BMNH collection, leg. L.M. Emms), and aphids collected on Arabis roots in England (BMNH collection, leg. G. Fox-Wilson) may also be this species. 2n=20.
Pemphigus populivenae Fitch Galls are elongate pocket-shaped or cock’s comb-shaped, greenish or yellowish or pale red, arising alongside mid-rib on upper side of leaf, with a slit-shaped opening underneath. On heavily-infested trees, galls may occur on both sides of the leaf, and those on the underside may then be confused with those of P. betae (Maxson & Knowlton 1934; however see also Footit et al. 2010). Whitham (1978) reported that P. populivenae galling Populus fremontii in Utah caused chlorosis of the leaf distal to the gall. As in the case of P. betae, several species of cottonwoods and their hybrids are colonised (angustifolia, balsamifera, deltoides, fremontii and trichocarpa; the latter species in experimental conditions only). The fundatrix is yellowish green. Alatae (BL 1.1-1.8 mm) emerge in June-July and migrate to found colonies on roots of Amaranthaceae. Rumex (Polygonaceae) may also be colonised. The evidence for this host alternation is work in California by Grigorick & Lange (1962), who transferred sexuparae from sugar beet to poplar bark under controlled conditions and obtained fundatrices that produced galls specifically on P. trichocarpa. The alatae from these galls were identified as P. populivenae, and colonised roots of B. vulgaris and Chenopodium album. However, the suggested synonymy of both P. betae and P. balsamiferae with populivenae indicates some confusion of identity, and further work seems advisable to confirm that the correct name for populations damaging sugar beet in California (e.g. Summers & Newton 1989) is in fact populivenae rather than betae. Foottit et al. (2010) have found molecular evidence that at least three taxa with similar galls occur sympatrically on cottonwoods in Alberta.
Pemphigus protospirae Lichtenstein Galls on Populus spp., especially P. nigra var. italica, are smooth, shiny, green mottled with red, caused by swelling, flattening and spiral twisting of the leaf petiole, similar to those of P. spyrothecae but often thinner and with more spirals (see fig. 131R and influential points/Gallery). P. protospirae, however, is heteroecious, and the alatae leave the gall in late May-early July to found colonies usually in the sheathing leaf-bases of aquatic Apiaceae (Berula erecta, Sium latifolium, Apium nodiflorum, Oenanthe aquatica). Apterae on secondary hosts are pale yellowish green or yellow, with abundant white wax especially on posterior abdomen; BL 1.6-2.3 mm. The return to Populus occurs in September-October. Widely distributed in Europe, but probably restricted by availability of secondary host habitats. Also recorded from west Siberia, Iran, Central Asia, eastern Siberia (where it is particularly asociated with P. laurifolia according to Babichev & Kirichenko 2020) and China (Zhang & Zhang 1979c).
Pemphigus rubiradicis Theobald Apterae are dirty greenish brown, with flocculent white wax on abdomen; BL 1.7-2.2 mm. On roots of Rubus idaeus in England (Kent).
Pemphigus saliciradicis (Börner) Apterae are yellowish white, lightly coated with greyish wax; BL 1.6-2.4 mm. On roots of Salix spp. (fragilis, herbacea, polaris) in Europe (records are from England, Finland, Sweden, Switzerland), Greenland (as salicicola Hille Ris Lambers), Transcaucasia (Georgia; but see below), north-east Russia (Wrangel Island; Khruleva & Stekolshchikov 2019) and Japan. Aphids on Salix roots in California, USA, and on Baffin Island in arctic Canada, are possibly also this species (but see below). Palaearctic populations identified as P. saliciradicis are apparently normally anholocyclic on Salix roots. A dense colony identified as this species was found on grapevine (Vitis vinifera) roots in Georgia (Barjadze 2010), but a subsequent morphometric study (Barjadze & Asanidze 2013) revealed differences from the willow root-feeding aphid (and see note below). Alatae that are intermediates between virginoparae and sexuparae occur in Japan (Aoki 1975). Lange (1965) reported Pemphigus populations of the saliciradicis group on Salix roots in northern and southern California; the southern population occurred throughout the year on Salix roots, whereas the the northern population produced sexuparae in autumn, but no descriptions were published of these aphids. Richards’ (1963) record of P. groenlandicus (a grass-root feeder) from roots of Salix reticulata on Baffin Island may be referable to this species.
[Note: There is a strong possibility that the aphids from willow roots in Europe and Asia, described as P. saliciradicis by Börner, are the introduced secondary host populations of the North American poplar-galling aphid Thecabius populimonilis, which is believed to have a partial migration to Salix roots; Aoki et al. (1996) noted a close resemblance between first instar nymphs of the two species, and data for apterous exules of T. populimonilis from Salix roots and mosses in Washington and Alaska, confirmed by DNA barcoding as this species (Pike et al. 2012), agree closely with P. saliciradicis (S. Bardjadze, pers. comm. 2013). If this is the case then the grapevine-feeding aphid found in Georgia will require a new name. The identity of a Pemphigus sp. collected from grapevine roots in Queensland, Australia, shown by Barjadze & Asanidze (2013) to differ morphologically from the grapevine root feeder in Georgia, will also need to be re-investigated.]
Pemphigus sinobursarius Zhang Galls on Populus simonii in China are on undersides of leaves at base of the lamina (Zhang & Zhang 1979c). Alatae similar to P. bursarius, but with a shorter R IV+V (0.065 mm). Migration occurs to an unknown secondary host.
Pemphigus siphunculatus Hille Ris Lambers Galls on twigs of Populus ciliata in Pakistan in August contained apterous progeny of the fundatrix, colour in life unknown (presumably wax-covered); BL 1.9-3.4 mm (Hille Ris Lambers 1973b). Presence of apterae in the gall is normally indicative of a monoecious life cycle but, according to S. Chakrabarti (pers. comm. cited by Aoki & Kurosu 1991d), P. siphunculatus is heteroecious. The secondary host is not recorded.
Pemphigus spyrothecae Passerini Gall is green, reddish or yellowish, smooth, formed by thickening, flattening and spiral twisting of petiole of Populus nigra (fig. 131S). Gall formation was studied by Dunn (1960). The fundatrix is pale green, giving rise to second generation apterae within the gall. Monoecious holocyclic; alate sexuparae emerge in August-November, to produce sexuales on the bark. Lampel (1960) provided a detailed account. First instars with thick forelegs function as soldiers and defend the gall against predators (Aoki & Kurosu 1986, Foster 1990), also repairing the gall (Pike & Foster 2004) and removing wax-coated droplets of honeydew (N. Pike et al. 2002). Urban (2002) made detailed studies of gall populations and the impact of natural enemies in the Czech Republic. N. Pike (2007) showed that different morphs occupy different positions within the gall, and N. Pike (2010) reported on the amino acid profile of this species. Widely distributed in Europe, in North Africa (Tunisia), Iran, Kazakhstan, Pakistan, eastern Siberia (Babichev & Kirichenko 2020), and introduced into Canada (Quebec, Alleyne & Morrison 1974; British Columbia). 2n=20.
Pemphigus tartareus Hottes & Frison (= Pemphigus junctisensoriatus Maxson) Described originally from alate sexuparae collected on roots of Bidens sp. in Illinois, USA. Galls on Populus deltoides are pale yellowish green, composed mostly of swollen leaf petiole around broadened base of leaf lamina, with an exit hole in the form of a long slit between the petiole and the leaf (Maxson 1934). The fundatrix is pale yellowish white, with brownish head and tibiae. Alatae (BL 1.4-1.8 mm) emerge in June-July and migrate to Bidens sp(p). Smith (1985) established the synonymy with P. junctisensoriatus and made successful host transfers from Populus to Dichondra repens (Convolvulaceae) as well as to Bidens. In USA and eastern Canada.
Pemphigus tibetapolygoni Zhang Apterae are presumably with wax; BL c.2.5 mm. On roots of a Polygonum sp. in Tibet.
Pemphigus tibetensis Zhang Galls are globular, pouch-like, formed on twigs of Populus cathayana in Tibet (Zhang & Zhong 1979c). This appears to be a member of the borealis/dorocola group.
Pemphigus trehernei Foster The gall induced experimentally on Populus nigra var. italica was green with reddish tinge around exit hole, globular, formed at base of current year’s shoot. The fundatrix is dull pale green. Alatae (BL 1.9-2.4 mm) emerge in June-July and found colonies on roots of Aster tripolium in salt-marsh habitats (Foster 1975), and probably specific to this halophytic host. Apterae on the secondary host are yellowish white to greenish white with greyish white wax forming tufts on abdomen; BL 1.3-2.4 mm. Alate sexuparae (with secondary rhinaria distributed III 3-7, IV 1-3) are produced in September-December. Anholocyclic overwintering on Aster roots is common, and the species has only been found naturally on its secondary host; the gall generations were obtained by experimental transfer of sexuparae to P. nigra. In England, Ireland and France.
Pemphigus turritus Zhang The galls on Populus purdomii in China (Gansu Prov.) are green, chilli- or horn-shaped, c. 15 mm × c. 3 mm thick. Described from three alatae (BL c.2.4 mm) collected in July (G. Zhang & Qiao 1997c). The life cycle is unknown.
Pemphigus venosus Habib & Ghani Recorded as forming galls on twigs of Populus ciliata in Pakistan, but no description is given, and therefore this is a nomen nudum.
Pemphigus (Pemphiginus) vesicarius Passerini Galls on Populus nigra originate from the mid-rib at the base of the upper side of the leaf, developing in April-May into an irregular pale green structure with numerous tubular outgrowths, of diameter up to 4 cm, that encompasses the whole leaf (fig. 131T and see influentialpoints.com/Gallery) . The fundatrix is dark slate-grey to blue-black. Alatae, BL 1.7-3.0 mm, leave the gall in May-June through secondary holes at apices of outgrowths (Roberti 1938) and migrate to found colonies on the stems and basal parts of Colutea arborescens. Apterae on Colutea (for long known as P. coluteae) secrete abundant wax and have BL 2.7-3.0 mm. Coffin & Remaudière (2001) determined the life cycle in southern France, described alate sexuparae and sexuales, and compared the various morphs with those of P. populi. The male genitalia were described and illustrated by Wieczorek et al. (2012). In southern Europe, Algeria, south-west and central Asia, east to Pakistan and India.
Pemphigus wuduensis Zhang The gall on Populus purdomii in China (Gansu Prov.) is flattened, rounded, and has a secondary exit. Described from two alatae (BL c. 2.4 mm) collected in July (G. Zhang & Qiao 1997c). The life cycle is unknown.
Pemphigus zhongi (Zhang) The gall on an unidentified Populus sp. is “dumpling-shaped”, formed by folding of the two halves of a leaf together, later becoming reddish, swollen and corrugated. Emigrant alatae are blackish green in life; BL c.2.6 mm (G. Zhang et al. 1995, as Thecabius (Parathecabius) zhongi). In Gansu province, China. The host is identified in Zhang (1999, p. 69) as P. cathayana. Except for the high rhinarial numbers and different gall, the alatae seem similar to those of P. matsumurai.
|
Pentalonia Coquerel |
Aphidinae: Macrosiphini |
Four species of small brownish aphids with symmetrically swollen siphunculi, the alatae having broadly dark-banded forewings with very characteristic venation, the radius strongly curved and almost touching or fused with the media for part of its length, making a closed or nearly closed cell (see aphids of Karnataka website). Foottit & Maw (2019) reviewed geographical distribution, host preferences and molecular diversity within the genus.
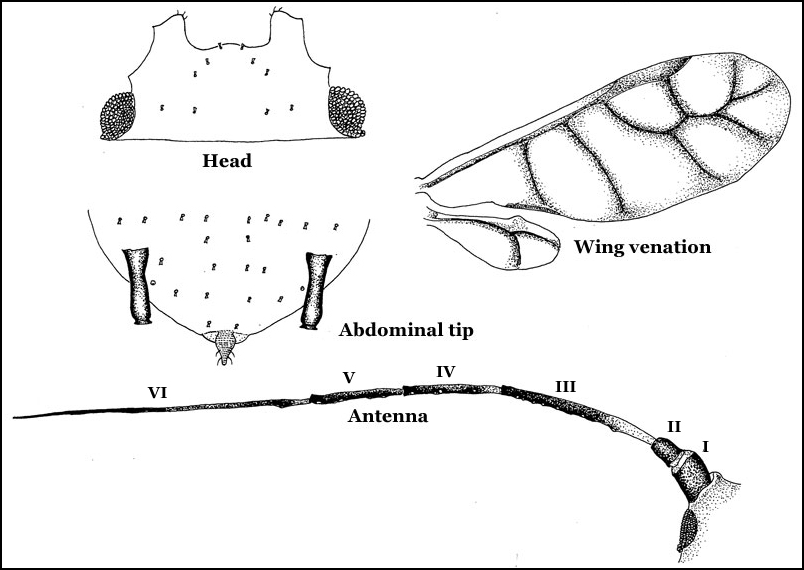{kind=link}
Pentalonia caladii van der Goot Apterae are reddish brown to almost black, with dark-tipped antennae; BL 1.0-1.7 mm. On plants in the families Zingiberaceae and Araceae (e.g. Alpinia, Caladium, Colocasia, Eletteria, Hedychium, Zingiber), or occasionally on plants in other families (Musa, Heliconius). Described from Java, distributed widely through the south Pacific, Hong Kong, Australia, Central America, and in hothouses in the northern hemisphere. This species has been long regarded as a synonym of P. nigronervosa, or as a “form” of that species, but molecular and multivariate studies by Foottit et al. (2010b) have shown that it should be treated as a distinct species. Bhadra & Agarwala (2010) compared fitness parameters of P. caladii and P. nigronervosa in reciprocal transfers between their respective host plants.
Pentalonia gavarri Eastop Apterae are dull black; BL 0.7-1.0 mm. Inside unfurling new grass blades (Centrotheca, Oplismenus). P. gavarri was originally described from alatae trapped in Philippines. Apterae described by Martin (1987) from West Malaysia, and specimens presumed to have been collected from Brachiaria sp. (although the host-plant is not explicitly stated) in India (Assam), described by A.K. Ghosh (1978) as Micromyzus brachiariae,clearly correspond well with this species. Alatae have also been trapped in New Britain and Australia (Carver & Hales 1983).
Pentalonia kalimpongensis (A.N. Basu) Apterae are reddish brown, with dark-tipped appendages; BL 1.5-1.9 mm. On Zingiberaceae (Curcuma, Elleteria, Hedychium) in India. In transfer experiments this species did well on other Zingiberaceae (Canna sp., Amomum sabulatum;original description, as Micromyzus). Monoecious holocyclic; oviparae and alate males were described from Elleteria by Mondal et al. (1978). 2n=12.
Pentalonia nigronervosa Coquerel Banana Aphid Plate 19f Apterae are reddish‑brown to almost black, with black-tipped antennae, and legs pale except for distal parts of femora (see aphids of Karnataka website and influentialpoints.com/Gallery); BL 1.2-1.9 mm. P. nigronervosa feeds almost entirely on plants in the family Musaceae, with a few records from Heliconiaceae and Zingiberaceae (Foottit & Maw 2019). It is often found living under the old leaf bases, usually ant-attended. It is an important vector of banana bunchy top virus (see also Blackman & Eastop 2000). P. nigronervosa is widespread through all tropical and subtropical parts of the world, and in hothouses in Europe, Australia and North America. Probably it is anholocyclic almost everywhere; in the BMNH collection there is one alate male from Nepal (leg. K.C. Sharma). The sexual morphs recorded from Curcuma in India by Bhanotar & L.K. Ghosh (1969) were possibly P. kalimpongensis. The complete genome sequence has been determined (Mathers et al. 2020). 2n=14.
|
Pentamyzus Hille Ris Lambers |
Aphidinae: Macrosiphini |
Four species in southern South America and one in western USA, possibly most closely related to Carolinaia, with 5-segmented antennae in all morphs. Four of the five species are grass feeders, and one (P. tenuis) has strongly modified morphology which parallels that of unrelated grass-feeding aphids. Nieti Nafría et al. (2002) reviewed the genus.
Pentamyzus acaenae (Schouteden) Apterae are pale green; BL 1.0-2.5 mm. On leaves and shoots of Acaena splendens in southern Argentina (Tierra del Fuego) and Chile (Magallanes; Nieto Nafría et al. 2018). Monoecious holocyclic with an abbreviated life cycle, oviparae and apterous males appearing in January (Nieto Nafría et al. 2002b).
Pentamyzus falklandicus Hille Ris Lambers Colour of apterae in life is unknown; BL 2.4-3.0 mm. On Poa flabellata in the Falkland Islands.
Pentamyzus fueginus Nieto Nafría, Mier Durante & Ortego Apterae are reddish brown to greenish brown; BL 1.8-2.6 mm. On outer-facing surfaces of leaves of Alopecurus magellanicus and Hordeum comosum growing in moist situations. In southern Argentina (Tierra del Fuego). Monoecious holocyclic with an abbreviated life cycle, oviparae and apterous males occurring in January (Nieto Nafría et al. 2002b).
Pentamyzus graminis Hille Ris Lambers Plate 12b Apterae are uniformly green; BL 1.6-2.0 mm. On basal parts of unidentified grasses, in somewhat moist and sunless situations, in California, USA. Monoecious holocyclic, with numerous oviparae and some apterous males occurring with apterous viviparae in March.
Pentamyzus tenuis Brown Colour of apterae in life is unknown, probably wax-covered; BL 2.3-2.9 mm. On Poa alopecurus on Falkland Islands, and subsequently found on Alopecurus magellanicus on Tierra del Fuego (Nieto Nafría et al. 2002b).
|
Periphyllus van der Hoeven |
Chaitophorinae: Chaitophorini |
About 50 species of medium-sized to large, generally long-haired aphids, almost all living on Acer except for four species on Aesculus and three on Koelreuteria. The antennal PT is usually long and the siphunculi are stump-shaped, usually with polygonal reticulation, but often longer than in Chaitophorus and with a pronounced flange. The cauda is either rounded or tongue-shaped with a slight constriction. Colonies are usually ant-attended. There is no host alternation, and the association with the host plant is an intimate one, involving a complex seasonal polymorphism to cope with physiological changes in the host, the most unusual feature of which is the frequent occurrence of specialised larval aestivating morphs (fig. 5). Periphyllus is predominantly a palaearctic genus, (15 European, 11 north Indian/Himalayan, 15 east Asian), with just 3 species native to North America. Essig & Abernathy (1952) wrote a monograph of the genus. There are keys and/or accounts of Periphyllus species of Europe (Hille Ris Lambers 1967a), Britain (Stroyan 1977), Fennoscandia and Denmark (Heie 1982), Iberian peninsula (Nieto Nafría & Mier Durante 1998), Poland (Wieczorek & Junkiert 2012), European Russia (Mamontova 1955), India (Chakrabarti et al. 1987), Pakistan (Remaudière 2002), China (Tao 1964, Li et al. 2022), east Siberia (Pashtshenko 1987a, 1988a, b), east Asia (Park & Park 1995), Japan (Sorin 1990, Sugimoto 2024) and Canada (Richards 1972a). Wieczorek & Junkiert (2011) and Trela et al. (2020) provided keys to oviparae and males of species found respectively in Europe and the Nearctic, and Lubiarz & Mackoś-Iwaszko (2015) provided morphometric data and keys to fundatrices of Periphyllus species living in Poland on Acer platanoides. Sugimoto (2024) revised the species found in Japan and provided a key to apterous morphs. Accounts of British species with excellent colour photographs are available online at http://influentialpoints.com/Gallery/Periphyllus_aphids.htm.
T. Liu et al. ( 2022) studied macroevolution of Periphyllus and other Chaitophorinae in relation to changes in host use. Recent molecular work indicates that Periphyllus is sister group to a clade containing Chaitophorus and Siphini (Wieczorek & Kajtoch 2011, Wieczorek et al. 2017b), but more recent results (T. Liu et al. 2022) suggest that the transition of Siphini to grasses may have occurred much earlier. Aphidius aerolatus, A. setiger, Praon negundinis and Trioxys falcatus are specialised parasitoids.
Periphyllus acericola (Walker) Apterae are pale green or yellowish green, sometimes with dorsal brownish markings; BL 2.4-3.5 mm. Alatae have broad dark dorsal abdominal cross-bars scarcely separated between segments, and paler marginal sclerites (see influentialpoints.com/Gallery); BL 3.0-3.5 mm. On undersides of leaves, petioles and young shoots of Acer pseudoplatanus throughout most of Europe, and usually regarded as specific to this host, but Lubiarz & Mackoś-Iwaszko (2015) recorded fundatrices on A. platanoides in Poland. Aestivating first instar nymphs are yellowish white with long, pointed dorsal and marginal hairs; they aggregate in dense groups appearing like whitish spots on undersides of leaves. Oviparae and alate males occur in October. 2n=18*.
Periphyllus acerihabitans Zhang Apterae are pale green; BL 1.9-2.5 mm. The antennal terminal process is unusually short, less than twice the length of the base of ANT VI. Alatae are without dark dorsal abdominal sclerotisation. On undersides of leaves, petioles and young shoots of Acer buergerianum, usually ant-attended. Aestivating nymphs have foliate marginal hairs. Oviparae occur in October-November, and were described by Wieczorek et al. (2016a), who also provided redescriptions of the apterous and alate viviparae. Males and other morphs are described in Sugimoto 2024. In China and Japan. Niu et al. (2016) compared the mitochondrial genome of P. acerihabitans with that of Chaetosiphella stipae.
Periphyllus acerijaponicus Sugimoto Apterae are pale yellow (spring and autumn generations); BL 1.57-2.6 mm (spring) and 2.6-3.0 mm (autumn). Alatae are yellowish-green with black bands dorsally (spring) or dark brown (autumn); BL 2.2-3.0 mm and 2.55-3.64 mm. On Acer japonicum from Japan (Sugimoto 2024).
Periphyllus aceriphaga Chakrabarti & Mandal Appearance in life is unknown; BL 2.4-2.7 mm. On Acer spp. (caesium, caudatum) in Uttar Pradesh, India (Chakrabarti et al. 1987), and Pakistan. Aestivating nymphs have foliate marginal hairs. Remaudière (2002) recorded oviparae and alate males collected from Acer sp. at Murree, Pakistan in November.
Periphyllus aceris (Linnaeus) Apterae are yellow, often with green flecks dorsally (see influentialpoints.com/Gallery); BL 1.5-3.7 mm. Alatae have dorsal cross-bands more widely separated than in P. acericola, with equally dark marginal sclerites and pterostigma; BL 3.2-4.5 mm. On undersides of leaves, petioles and growing shoots of Acer spp., especially A. platanoides, which is clearly its normal host. There are also records from Aesculus, but no authentic specimens have been seen and these should possibly be referred to P. testudinaceus. Throughout most of Europe, and also recorded from Iran (Abaii 1984). Previously stated to have been introduced to North America, but Foottit et al. (2006) point out that all North American specimens in USNM are interceptions. Aestivating nymphs have long pointed dorsal and marginal hairs and aggregate like those of P. acericola. Monoecious holocyclic; oviparae and alate males occur in October-November. The fundatrix was described by Lubiarz & Mackoś-Iwaszko (2015). This species has been confused in the literature with acericola, and also with other species; e.g. Börner (1940) had P. lyropictus, and Wahlgren (1935) had P. testudinaceus. 2n=16 (Blackman & Eastop 1994).
Periphyllus aesculi Hille Ris Lambers Apterae are rather elongate oval, brownish in life with black dorsal spots (A.K. Ghosh 1980); BL 2.5-3.5 mm. Siphunculi are wholly dark. Alatae have short dorsal abdominal cross-bars and dark marginal sclerites. On undersides of young leaves of Aesculus indicus in India (Kashmir, Himachal Pradesh, Uttar Pradesh) and Pakistan (Murree; Remaudière 2002). Chakrabarti et al. (1972a) described the apterous vivipara and redescribed the alate vivipara, L.K. Ghosh (1986) described the ovipara, and Agarwala & Raychaudhuri (1982) described the alate male. Aestivating morphs are not recorded. Sexuales were collected in October-December.
Periphyllus allogenes Szelegiewicz Apterae are elongate oval, brown with dark brown antennae and legs; BL 1.0-1.2 mm. Alatae are unknown. On leaf petioles of Acer triflorum in Korea and Japan (Sugimoto 2024). Described from autumnal (late August) apterae, which may have developed from aestivating nymphs and be very different from spring forms, which are possibly much larger (Szelegiewicz 1981b). However apterae of P. triflorumi, also described from A. triflorum in Korea without reference to P. allogenes and presumably a synonym, are equally small and were collected in July (Lee & Seo 1992). Differences in specimens from Japan and Korea are described in Sugimoto (2024). The biology is unknown.
Periphyllus americanus Baker Apterae in life are “pale yellowish green with dark median stripe on abdomen forming a cross in region of cornicles” (Hottes, quoted by Palmer, 1952); BL 2.0-2.5 mm. The alata is dark greenish with black dorsal abdominal cross-bars and marginal sclerites. On leaves of Acer spp., especially sugar maples, widely distributed in North America. Aestivating nymphs have foliate marginal hairs (fig. 19A). Essig & Abernathy (1952) gave an account of the summer generations. Fundatrices and oviparae have apparently not been described. Trela et al. (2020) described an alate male (leg. G.F. Knowlton), collected in Utah in October.
Periphyllus bengalensis Ghosh & Raychaudhuri Apterae are yellow, with dark siphunculi, pear-shaped; BL 1.8-2.2 mm. Alatae have dark dorsal abdominal cross-bands. Forming a colony on undersides of older leaves of an undentified Acer sp. in West Bengal, India (A.K. Ghosh 1980). Aestivating and sexual morphs are unknown.
Periphyllus amamiensis Sugimoto   Apterae are pale green (spring) or dark green to dark brown(autumn); BL 1.9-2.6 (spring) or 2.4-3.3 (autumn) mm. Alatae are pale green with black bands dorsally; BL 2.6-3.3 mm. Other morphs have been described. On Acer insulare in Japan (Sugimoto 2024).
Periphyllus blackmani Li & Huang   Apterae are dark green with wholly black tibiae and siphunculi, and often with paired dark spinal and marginal spots on abdominal segments. BL 2.2-3.6 mm. Alatae have dark dorsal abdominal cross-bars and marginal spots, and 32-48 secondary rhinaria on ANT III. On Koelreuteria bipinnata and K. paniculate in subtropical humid areas of eastern and southeastern China. No data are known on aestivating nymphs.
Periphyllus brevisetosus Sorin Appearance in life is unknown; only late summer apterae (BL 1.2-1.3 mm) are described, collected in Japan on Acer pictum var. typicum (= mono) (Sorin 1990b). Closely related to P. kuwanaii.
Periphyllus brevispinosus Gillette & Palmer Apterae are brown with dark and pale mottling, and pale siphunculi; BL 2.4-2.9 mm. Alatae are brown with black dorsal abdomnal cross-bands and marginal sclerites. In undersides of leaves of Acer glabrum in western North America. Aestivating nymphs have foliate marginal hairs with serrate margins (Essig & Abernathy 1952). Monoecious holocyclic; ovipara and alate males collected in mid September were described by Trela et al. (2020).
Periphyllus bulgaricus Tashev Apterae are shining black, with pale legs and antennae; BL 2.0-2.8 mm. Apterae and alatae both have variably-developed black dorsal abdominal cross-bars (see Nieto Nafría & Mier Durante 1998). On shoot tips and undersides of leaves, usually at base of mid-rib. Described from Acer hyrcanum in Bulgaria (Tashev 1964c); also collected from A, hyrcanum in Turkey, from A. opalus and A. ibericum in Iran, from A. hermaneum in Lebanon (BMNH collection, leg. D. Hille Ris Lambers), and from A. granatense and A. monspessulanum in Spain. Nieto Nafria & Gonzales Funes (1990) described fundatrices, oviparae and alate males, and discussed the very difficult separation of this species from P. rhenanus. Aestivating nymphs have not been recorded.
Periphyllus caesium Chakrabarti & Saha Appearance in life is not recorded, probably rather pale with dark siphunculi; BL of aptera 2.3-2.4 mm. Alatae have dark dorsal abdominal cross-bands. On Acer caesium in Uttar Pradesh, India (Chakrabarti et al. 1987). An alate male ascribed to this species was collected in October. Longer haired than P. aceriphaga, and with a shorter HT II, but otherwise apparently very similar.
Periphyllus californiensis (Shinji) Apterae are dark olive-green to brown, with dark brown head and prothorax, and irregular dark dorsal segmental spots or cross-bands (see influentialpoints.com/Gallery); BL 2.3-3.5 mm. Alatae have dorsal abdominal cross-bands which with a hand lens appear darker than the pterostigma of the forewing (cf. aceris). Additional diagnostic characters have been added by Sugimoto (2024). On Acer spp. in east Asia (China, Japan, east Siberia, Korea, Taiwan, Thailand, West Bengal), living on undersides of leaves along veins; Pashtshenko (1987) gave A. pseudosieboldianum as the principal food plant. Also recorded from Aesculus turbinata in Japan (as Chaitophorinella acerifoliae; Takahashi 1919a). Introduced to Europe, North America, Australia and New Zealand, where it is most commonly found on planted Asian ornamental maples, but sometimes also colonises native species; there is one record from Aesculus californica in California. Aestivating nymphs have foliate marginal hairs. Essig & Abernathy (1952) fully described the life cycle in California, USA, where oviparae and alate males occur in October-November. Population dynamics of P. californiensis in Japan has been studied in relation to host phenology (Hashimoto & Furuta 1988, Wang 2006), mortality of aestivating nymphs (Furuta 1985), host preferences (Furuta 1986) and natural enemies (Furuta & Mao 1986, Kan & Sasakawa 1986). 2n=18 (Blackman 1986).
Periphyllus changlai Remaudière Apterae are pale-coloured, long-bodied; BL 3.2-3.8 mm. Alatae are without dorsal abdominal sclerotisation. On Acer caesium in Pakistan (Remaudière 2002). An ovipara and alate males were collected at the same time as the parthenogenetic morphs in October.
Periphyllus chidoricola Sugimoto Apterae are greenish black with larvae pale green (spring) or dark brown with hind femora and base of hind tibiae brownish (autumn); BL 1.3-1.8 (spring) or 1.9-3.1 (autumn) mm. Alatae are pale green to green with black bands dorsally; BL 1.9-3.4 mm. Other morphs have been described. In Japan on Acer carpinifolium (Sugimoto 2024).
Periphyllus coracinus (Koch) Apterae are variable in colour, yellow, light or dark green, or brown, or mottled with green or brown. On leaf petioles and twigs of Acer platanoidis in western, central and eastern Europe (Netherlands, Sweden, Finland, Germany, Austria, Poland, Hungary, Moldova, Ukraine, Turkey), and also recorded from Kazakhstan (Kadyrbekov 2017a). Oviparae and alate males occur in October. The male genitalia were described and illustrated by Wieczorek et al. (2011). The fundatrix was described by Lubiarz & Mackoś-Iwaszko (2015). Apparently no specialised aestivating nymphs are produced.
Periphyllus formosanus Takahashi Apterae are reddish brown, with thorax darker; BL c. 2.5 mm. Alatae have blackish dorsal abdominal cross-bands. On undersides of leaves of unidentified Acer sp(p). in Taiwan and China. Aestivating nymphs have foliate marginal hairs (Takahashi 1933a). Probable fundatrices were observed by Takahashi in February, and Essig & Abernathy (1952) described the oviparae and alate males, collected in November.
Periphyllus garwhalensis Chakrabarti & Mandal Apterae are undescribed. Alatae (BL 3.0-3.6 mm) have dark siphunculi but no dorsal abdominal cross-bands. On Acer caudatum in Uttar Pradesh. India. Aestivating nymphs have foliate marginal hairs with serrate margins (Chakrabarti et al. 1987).
Periphyllus ginnalae Paik ex Szelegiewicz Apterae and alatae are shining dark brown to black; BL 1.6-2.6 mm. Alatae have secondary rhinaria distributed ANT III 10-19, IV 0-4, V 0-2. On Acer ginnala, living in early spring on leaf petioles at tips of young branches, and later forming dense colonies on the undersides of the leaves. Aestivating nymphs are green to brown with foliate marginal hairs, living singly on both sides of leaves; but most of the population apparently does not aestivate (Pashtshenko 1988b, as loricatus). Apterous males and oviparae occur in September (Pashtshenko 1987a). In Korea and east Siberia, and possibly also in Japan, as P. takahashii Sorin (1990b), described from A. ginnala in Japan, seem likely to be small specimens of this species. [Park & Park (1995) questioned the synonymy of ginnalae, loricatus and takahashii proposed in Blackman & Eastop (1994). Although they appear to be correct in claiming validity for the name ginnalae, which had been listed as a nomen nudum (Eastop & Hille Ris Lambers 1976, Remaudière & Remaudière 1997), specimens in the BMNH collection from A. ginnala in both North and South Korea determined as ginnalae by Paik and by Szelegiewicz agree well with the description of P. loricatus (Pashtshenko 1987), in particular having 7 hairs on first tarsal segments and R IV+V 0.7-0.8 × HT II.] Sugimoto (2024) has treated P. loricatus as a valid species and provided a redescription.
Periphyllus guangxuei Li & Huang Apterae are light green, with relatively small dark brown dorsal spots on thorax and abdomen, front and middle tibiae most pale and hind tibiae apically pigmented. BL 2.0-2.7 mm. Alatae have dark dorsal abdominal cross-bars and marginal spots, most part of hind leg dark brown and 29-37 secondary rhinaria on ANT III. On Koelreuteria bipinnata in the subtropical highland region in Yunnan (China). No data are known on aestivating nymphs.
Periphyllus himalayensis Chakrabarti Apterae are undescribed. Alatae have dark dorsal abdominal cross-bands, colour in life unknown, BL 3.1-3.5 mm (Chakrabarti 1977). On unidentified Acer sp(p). in northern India (Himachal Pradesh, West Bengal). Biology is unknown. Perhaps these are large specimens of P. bengalensis.
Periphyllus hirticornis (Walker) (fig. 23B, fig. 25A) Apterae are light green, with red eyes, without any dark dorsal markings (see influentialpoints.com/Gallery); BL 2.0-3.0 mm. The abdomen of the alata has variably developed spinal sclerites, but not cross-bands. On undersides of leaves, leaf petioles and fruits of Acer campestre, often with attendant ants. Aestivating nymphs have foliate marginal hairs (see Stroyan 1977). Widely distributed in Europe. Oviparae and alate males are produced in October-November. 2n=18*.
Periphyllus hokkaidensis Sorin Apterae are dark reddish to blackish brown; BL c. 2 mm. On Acer miyabei and A. mono var. subtrifidum in Hokkaido, Japan (Sorin 1990b). Oviparous females, males and additional characters of apterae are described in Sugimoto (2024). Other morphs and biology are unknown. It seems to be closely related to P. kuwanaii.
Periphyllus kajicola Sugimoto Apterae are shining black; BL 1.8-3.4 mm. Alatae have a dark brown abdomen with black bands dorsally; BL 2.3-3.4 mm. Other morphs have been described. Mature dimorphs might produce oviparous females directly (Sugimoto 2024). On Acer diabolicum in Japan.
Periphyllus karatavicus Kadyrbekov Apterae pale green; BL c.1.5-1.6 mm. On upper and undersides of young leaves of Acer semenovii in Kazakhstan (Kadyrbekov 1999b).
Periphyllus koelreuteriae (Takahashi) Apterae are variably pigmented, yellow or black, with wholly black tibiae and siphunculi, and often with paired dark spinal and marginal spots on abdominal segments; BL 1.6-3.3 mm. Alatae have dark dorsal abdominal cross-bars and marginal spots, and 25-42 secondary rhinaria on ANT III. On Koelreuteria paniculata in in temperate areas of northern China, Korea and Japan. Aestivating nymphs have foliate marginal hairs (Takahashi 1933a, Essig & Abernathy 1952). The life cycle in Taiwan was studied by Y.C. Liu et al. (1999), the sexual morphs were described by Junkiert & Wieczorek (2019) and full data of apterous and alate viviparae were provided by Li et al. 2020. The recorded occurrence on Aesculus turbinata based on a single alata (Higuchi 1972) is casual (Li et al. 2022). Sorin’s (1990b) apterae from A turbinata keyed as P. koelreuteriae do not seem to be this species. 2n=18 (Blackman & Eastop 1994).
Periphyllus kuwanaii (Takahashi) Apterae and alatae are shining black; BL 1.4-2.4 mm. Principally on Acer mono; Pashtshenko (1988 a,b) recorded it from several other Acer spp., but such visits may be casual. In China, east Siberia, Japan and Korea. Aestivating nymphs have foliate marginal hairs (Takahashi 1933a); but part of the population does not aestivate (Pashtshenko 1988b). Sexual morphs are apparently undescribed. Periphyllus diacerivorus Zhang (in Zhang & Zhong 1982c) seems to be a synonym. P. tokyoensis have been designated as a synonym (Sugimoto 2024). 2n=18 (Blackman & Eastop 1994).
Periphyllus loricatus Pashtshenko Apterae have a head, thorax, and spinal area of abdomen shiny black, the rest of abdomen dark brown; BL 1.7-2.6 mm. Alatae almost black; BL 2.18-2.3 mm. On Acer ginnala in Japan, the Korean Peninsula and the Russian Far East (Sugimoto 2024). Previously considered a synonym of P. ginnalae but Sugimoto (2024) treated it as a valid species and gave redescriptions of fundatrices, apterae (spring and summer generations) and alatae.
Periphyllus lyropictus (Kessler) (fig. 23D) Apterae are yellowish with variably developed brown or green dorsal markings, usually comprising a broad spinal stripe on head and thorax and a large V-shaped mark on the dorsal abdomen (see influentialpoints.com/Gallery); BL 1.9-3.0 mm. Alatae have dark marginal sclerites, but other dorsal sclerotisation is limited to the spinal area, not forming cross-bands. On undersides of leaves of Acer platanoides, often forming large colonies producing much honeydew and visited by ants and other insects. This species does not produce aestivating nymphs. In Europe, Iran (Rezwani et al. 1994), and introduced on Norway maple to North America, where it is also widespread. A. platanoides is clearly the favoured host, and records from other Acer are mostly due to confusion with P. aceris and P. testudinaceus, particularly in the North American literature (see Essig & Abernathy 1952). Oviparae and alate males are produced in October-November.
Periphyllus mamontovae Narzikulov Apterae are green; BL 2.1-2.9 mm. On leaves of Acer turkestanicum and (more rarely) A. regelii in Tajikistan (Narzikulov 1957). Aestivating nymphs are apparently not produced. Life cycle and sexuales have not been described.
Periphyllus mandshuricus Pashtshenko Apterae are apparently not produced in spring; reddish brown to dark brown fundatrices on shoot tips produce only dark brown alatae (BL 2.5-3.4 mm), which live on undersides of leaves and produce aestivating nymphs with foliate marginal hairs. On Acer mandshuricum in maritime region of east Siberia. Oviparae and alate males ocur in October (Pashtshenko 1988b).
Periphyllus minutus Shaposhnikov Apterae are shining brown or greenish yellow, with dark brown dorsal markings; BL 1.6-2.5 mm. Alatae have dark dorsal abdominal cross-bands. On Acer tataricum, colonising young shoots, leaf petioles, undersides of leaves, fruit stalks and fruits. This species is also recorded from A. ginnala (Ripka et al. 1998) and A. campestre (Vereshchagin et al. 1985). Aestivating nymphs have long pointed dorsal and marginal hairs. In eastern Europe, south-west and central Asia (Bulgaria, Hungary, Moldovia, Ukraine, Iran, Kazakhstan). Sexuales have not been described. Described as a subspecies of aceris by Shaposhnikov (1952); Mamontova (1955) provided a fuller description and gave it full species status.
Periphyllus negundinis (Thomas) Apterae are pale yellow-green to apple green with dark dorsal markings (see influentialpoints.com/Gallery); BL 2.0-2.5 mm. Alatae have variable dark green dorsal abdominal markings, but no distinct cross-bands. On Acer negundo, feeding on young growth in or near fruit clusters in spring, then on undersides of leaves. Widely distributed in North America, including Mexico. Aestivating nymphs have foliate marginal hairs. Apterous males and oviparae occur in late September to November. Essig & Abernathy (1952) provided detailed descriptions of the life cycle, and Trela et al. (2020) redescribed the sexual morphs. 2n=20.
Periphyllus nevskyii Mamontova Apterae are shining grey-black, with a central black sclerite extending across ABD TERG 4-6; BL 1.3-2.0 mm (Nevsky 1929a; as Chaitophorinella negundinis; Mamontova 1955). On Acer regelii in central Asia (Tajikistan, Kazakhstan, Uzbekistan). Also recorded from A. semenovii (Kadyrbekov 2002), A. turkestanica and A. pseudoplatanus (Gabrid 1989). The life cycle has not been studied.
Periphyllus obscurus Mamontova (fig. 23C, fig. 24B) Apterae are blackish green (see influentialpoints.com/Gallery); BL 1.8-2.6 mm. In ant-attended colonies on young shoots, leaf petioles and undersides of leaves of Acer campestre in Europe (Bulgaria, Czechoslovakia, England, Netherlands, Germany, Hungary, Poland, Turkey, Ukraine) and Iran (Abaii 1984). Aestivating nymphs are apparently not produced. Sexual morphs have not been described.
Periphyllus pakistanicus Remaudière Apterae pale-coloured with dark apices to antennal segments, dark siphunculi, and dark spots at bases of dorsal hairs; BL 2.1-2.5 mm. Alatae are without dorsal abdominal cross-bands. In spring populations on Acer oblongus in Pakistan (Remaudière 2002).
Periphyllus pallidus Chakrabarti & Saha Appearance in life not recorded; BL of aptera 2.4-2.7 mm. On an unidentified Acer sp. in Uttar Pradesh, India (Chakrabarti et al. 1987). Aestivating nymphs have foliate marginal hairs. Other morphs and life cycle are unknown.
Periphyllus rhenanus (Börner) Apterae are shining, dark; BL 2.5-3.0 mm. On undersides of leaves, extending onto petioles and twigs. Alatae have broad black dorsal abdominal cross-bars and equally dark marginal sclerites. On Acer monspessulanum and A. obtusatum in southern and central Europe (Bulgaria, France, Germany, Greece, Italy, Switzerland). Aestivating nymphs have very long marginal hairs and form dense clusters (Barbagallo & Stroyan 1982). Apterous males and oviparae occur in October-November (Hille Ris Lambers 1947d; as P. lichtensteini). Nieto Nafría & Gonzales Funes (1990) discussed differences from the closely-related P. bulgaricus.
Periphyllus singeri (Börner) Apterae are reddish brown, in spring bearing dark dorsal abdominal cross-bands like the alatae; BL 2.4-3.5 mm. On young shoots or branches of young trees of Acer pseudoplatanus, or on old wounds on the bark of large tree trunks, sometimes in large colonies, attended by ants (Lasius fuliginosus). In Europe (Austria, Denmark, Germany, Lithuania, Netherlands, Ukraine), but apparently rare or sporadic in occurrence. Aestivating nymphs are apparently not produced. Oviparae and alate males occur in October-November, and were fully described and illustrated by Wieczorek & Junkiert (2011). The male genitalia were described and illustrated by Wieczorek et al. (2011).
Periphyllus steveni Mamontova Apterae are elongate oval, shining brown or dark green; BL 1.8-2.4 mm. On young growth and leaves of Acer stevenii in the Crimea (Mamontova-Solukha 1962). One of the group of species with long black dorsal hairs (Nieto Nafria & Gonzales Funes 1990). Aestivating nymphs are yellow, with long pointed hairs. Life cycle and sexual morphs have not been described.
Periphyllus tegmentosus Pashtshenko The fundatrix is green with brown markings; her progeny are all alate, having shining greenish yellow abdomen with dark dorsal adbominal cross-bars and margunal sclerites. BL of alata 2.2-2.9 mm. On Acer tegmentosum in maritime east Siberia. Aestivating nymphs yellow-green with foliate marginal hairs. Oviparae and alate males occur in October (Pashtshenko 1988b).
Periphyllus testudinaceus (Ferni) Plate 12c, d (fig. 23A, fig. 24C) Apterae are dirty dark green to dark brown or blackish (see influentialpoints.com/Gallery); BL 2.0-3.7 mm. Alatae have dark dorsal abdominal cross-bands, darker than pterostigma, and equally pigmented marginal sclerites. On young growth, leaves and leaf petioles of numerous Acer spp. (but especially common on A. campestre), and sometimes on Aesculus spp. Throughout Europe, in Iran (Hodjat 1993), in Korea (needs confirmation), and introduced to Tasmania, New Zealand and North America – where it is widely distributed. Aestivating nymphs with foliate marginal hairs are the only morph occurring from June to September (fig. 5). Oviparae and alate males occur in October-November. Essig & Abernathy (1952) provided a detailed account of the life cycle and polymorphism. Warrington & Whittaker (1985) showed that ant attendance caused a large increase in sap ingestion by this species. 2n=18.
Periphyllus unmoonsanensis Park & Park Apterae are “generally brown”; BL 2.6-2.8 mm. Alatae are yellowish green. On Acer palmatum in Korea (Park & Park 1996).
Periphyllus vandenboschi Hille Ris Lambers Colour of aptera in life was not recorded but probably pale green, with contrastingly black siphunculi; BL 1.4-2.0 mm. Alata has pale brown abdomen with rather small and indistinct spinopleural sclerites on ABD TERG 1-6 and cross-bands on ABD TERG 7 and 8 (Remaudière 2002). Collected on leaf petioles of an unidentified Acer in Pakistan (Hille Ris Lambers 1966c), subsequently found on A. caesium and ornamental A. oblongum (Naumann-Etienne & Remaudière 1995), and also recorded from Himachal Pradesh, India (see L.K. Ghosh 1986). Aestivating nymphs and other morphs, including sexuales, have not been recorded. Chakrabarti et al. (1972a) had P. villosii.
Periphyllus venetianus Hille Ris Lambers (fig. 24A, fig. 25B) Apterae are shining blackish green with pale antennae and legs; BL 1.1-2.0 mm. Alatae are undescribed. On undersides of leaves and leaf petioles of Acer campestre, attended by ants. Aestivating nymphs have foliate marginal hairs. In southern Europe (Austria, Czechoslovakia, France, Italy, Yugoslavia). Barbagallo & Stroyan (1982) recorded immature oviparae in Sicily in September.
Periphyllus villosii Chakrabarti Aptera are elongate oval, large, probably pale; BL c. 3.5 mm. The abdomen of the alata is without dark dorsal cross-bands. On Acer spp. (caesium, caudatum, villosum) in Uttar Pradesh, India. Aestivating nymphs have foliate marginal hairs (Chakrabarti et al. 1987). Oviparae and alate males occur (on A. villosum) in October (Chakrabarti 1977). Remaudière (2002) reported this species from Acer caesium in Pakistan, but noted differences from the original description that might indicate the existence of two distinct species.
Periphyllus viridis (Matsumura) Apterae (?fundatrices) are green; BL 2.9-3.9 mm. Alatae (in spring) are without dorsal abdominal cross-bands; according to Pashtshenko (1987), all progeny of the fundatrix are alate. On young growth, leaves and leaf petioles of Acer mono in Japan, Korea and maritime east Siberia. Aestivating nymphs have foliate marginal hairs. Oviparae and alate males occur in October; Pashtshenko (1987) gave a full account of the life cycle. Descriptions of specimens from Japan are given in Sugimoto (2024). Sorin (1990b) described some large alate specimens in Japan as a subspecies, P. viridis osugiensis. New morphs of this subspecies are described in Sugimoto (2024).
|
Phloeomyzus Horvath |
Phloeomyzinae |
Perhaps only one species, unique in morphology and in that the only alatae are sexuales. Parthenogenetic forms are all apterous, with fused head and prothorax and 3-faceted eyes.
Phloeomyzus passerinii Signoret Plate 2e, f Apterae are green, covered with dirty white wax wool; BL 1.2-2.2 mm. On bark and in crevices on trunks of Populus spp., heaviest infestations occurring on 6 to 8-year-old trees. There is no host alternation. Oviparae and males are produced in September-October in the northern hemisphere, the oviparae each laying two eggs (Theobald 1929). The male genitalia were described and illustrated by Wieczorek et al. (2011). Anholocyclic overwintering by apterous viviparae is also common, and may predominate; no morph recognisable as a fundatrix has been described. In Europe, north Africa (Egypt, Morocco), south-west, central and east Asia (incl. Japan; BMNH collection, leg. S.Aoki); USA (only record is from Maine) and South America (Argentina, Chile, Colombia). Arzone & Vidano (1984) studied the life cycle in Italy. The European P. nigra-feeding form was described as a separate species, Ph. redelei Hille Ris Lambers 1931 (see Doon & Hille Ris Lambers 1962), as also were large specimens found on P. tremuloides in Maine, USA (as Ph. dearborni; Smith 1974b). Zhang (in Zhang & Zhong 1982) described a subspecies on P. lasiocarpa in China, P. passerinii zhangwuensis, which was redescribed by Qiao et al. (2018). It seems likely, however, that a single, somewhat variable species is involved. 2n=10.
|
Phorodon Passerini |
Aphidinae: Macrosiphini |
A small genus of Myzus-like species with finger-like processes on antennal tubercles and a secondary host association with Cannabaceae (Cannabis, Humulus). The host-alternating species have Prunus as primary hosts. See also Blackman & Eastop 2000, p. 321-2.
Phorodon (Diphorodon) cannabis Passerini Cannabis Aphid Plate 19h Apterae are shiny yellowish green with three green longitudinal stripes, or pale to mid-green or pale pink without stripes (see influentialpoints.com/Gallery); BL 1.5-2.7 mm. Alatae have a dark dorsal abdominal patch, and secondary rhinaria distributed III 15-30, IV 2-13, V 0-2. On undersides of leaves of and on flower stems of Cannabis spp. In central, eastern and southern Europe, across Asia to Japan, in North Africa, and now recognised to occur widely in North America (Cranshaw et al. 2018). Monoecious holocyclic, with oviparae and alate males in September-November (Müller & Karl 1976). 2n=12.
Phorodon humuli (Schrank) Damson-Hop Aphid Plate 19i Apterae on Prunus in spring are pale green to yellowish green, rather shiny, with three darker green dorsal longitudinal stripes (see influentialpoints.com/Gallery); BL 2.0-2.6 mm. Alatae have a black dorsal abdominal patch and secondary rhinaria distributed III 23-35, IV 1-11, V 0. Spring colonies occur on Prunus insititia and related species (domestica, mahaleb, spinosa), causing only slight leaf curl. It is a vector of plum pox virus. Migration occurs in May-June to hops (Humulus lupulus), on which its importance as a pest has increased with development of insecticide resistance (Hrdy et al. 1986). Apterae feed on undersides of leaves and on flowers and fruits of hops and are pale green with darker green medial and lateral longitudinal stripes; BL 1.0-2.1 mm. Return migration to Prunus (studied by Eppler 1988) is in September-October. In Europe, north Africa (Ethiopia), south-west Asia (Iran, Turkey), Central Asia; and introduced to North America, where it is widespread, and New Zealand (BMNH collection, leg. P.L. Robertson). Morphometric variation was studied by Hampson & Madge (1986 a,b), and allozyme variation by Eggers Schumacher & Sander (1988). The photoperiodic determination of sexual morphs was studied by Campbell & Tregidga (2005). See also Blackman & Eastop 2000, p.321). 2n=12.
Phorodon humulifoliae Tseng & Tao Apterae are pale yellowish green or green with dark head, ANT, legs, and cauda; BL c.1.8 mm. Siphunculi are strongly curved outward, and brownish towards their apices. On leaves of Humulus japonicus in China (Chekiang, Szechuan). Presumably there is host alternation to Prunus spp. as primary hosts, as there are records from north-west China of this species occurring on P. armeniaca and P. salicina (G. Zhang 1999). The record of P. humulifoliae from Ulmus sp. (Tao 1963) is probably in error. 2n=12 (Chen & Zhang, 1985b).
Phorodon japonensis Takahashi Apterae and alatae resemble those of P. humuli (Takahashi 1965c); BL 1.4-1.8 mm. On Prunus spp. (mume, salicina) in Japan, Korea, east Siberia and Taiwan. Migration occurs to Humulus spp. (japonicus, scandens). It was described as a subspecies of humuli, to which it is closely related. 2n=12.
Phorodon viburni Matsumura Apterae are undescribed, alatae are brownish, BL c.1.5 mm. On undersides of leaves of a Viburnum sp. in Japan. This species has apparently not been found since its original description in 1918.
|
Phyllaphis Koch |
Phyllaphidinae |
A genus with two or three species on Fagus with well-developed dorsal wax glands, a very short processus terminalis and pore-like siphunculi. Takahashi (1960) distinguished it from related genera with a key, and Stroyan (1977) provided a generic diagnosis. Quednau (2010) reviewed the genus and provided illustrations of all available morphs. Trioxys phyllaphidis is a specialised parasitoid.
Phyllaphis fagi (Linnaeus) Plate 8 e,f Apterae are elongate oval, pale yellowish green, covered with wax wool (see influentialpoints.com/Gallery); BL usually 2.0-3.2 mm (but summer dwarfs down to 1.1 mm). Alatae have abdomen wax-covered, with variably-developed dark dorsal cross-bars. On undersides of mostly young leaves of Fagus spp., causing them to curl downwards on both sides of the mid-rib, and often to wither and die prematurely. Throughout Europe, east to Turkey and Caucasus, more recently reported from China (Qiao & Zhang 2004a) and Korea (Quednau & Lee 2001), and introduced to Australia, New Zealand and North America. Records from Japan possibly all apply to P. fagifoliae. Monoecious holocyclic, with dwarf apterae living through summer months to produce apterous oviparae and alate males in October-November. Oviparae have variably developed dark dorsal cross-bars (cf. fagifoliae and grandifoliae). Wieczorek & Świątek (2008) studied morphology and ultrastructure of the male reproductive system. Chandler (1968) studied its association with a syrphid predator. 2n=16.
Phyllaphis fagifoliae Takahashi Apterae are pale, presumably with wax; BL 1.2-2.5 mm. Alatae have much less pigmentation than P. fagi (Takahashi 1937). On Fagus spp. (crenata, japonica) in Japan. A record from Turkey (Görür et al. 2011a) is doubtful and requires confirmation. Sexuales occur in October (Higuchi 1972; as P. fagi). Oviparae are pale without dorsal abdominal cross-bars (Quednau 2010). 2n=26 (after Shinji; see Blackman 1986). [This species was synonymised with P. fagi by Higuchi, but the difference in chromosome number coupled with morphological differences indicate that it is a distinct species.]
Phyllaphis grandifoliae Richards Apterae are pale greenish, pale yellow or somewhat cream-coloured (Richards 1973), usually exuding filamentous wax (see influentialpoints.com/Gallery); BL c. 1.8 mm. On Fagus grandifolia in western North America. Alatae have not been recorded. Oviparae have pale siphunculi and lack dorsal abdominal cross-bars (Smith & Parron 1978, Quednau 2010).
Phyllaphis nigra Ashmead Apterae are shiny black; BL c. 1.3 mm. On tender shoots of Quercus phellos var. laurifoliae (= Q. laurifolia) in Florida, USA (Ashmead 1881). It has not been recorded since its original description, and almost certainly does not belong in this genus; it is perhaps a Lachnochaitophorus. The life cycle is not known.
|
Phyllaphoides Takahashi |
Calaphidinae: Panaphidini |
A genus for one east Asian narrow-bodied species living on bamboos. Quednau (2003) illustrated the available morphs.
Phyllaphoides bambusicola Takahashi Alatae are white with pale yellow mesothorax, secreting white cottony wax (see Martin & Lau 2011 for photograph and influentialpoints.com/Gallery); BL 1.5-2.5 mm. Apterae are also reported to occur (Takahashi 1923), but do not seem to have been described. On undersides of leaves of Bambusa stenostachya (Takahashi 1923) and Phyllostachys spp. (Liao 1976) in Taiwan and China (Tao 1964, Martin & Lau 2011). Quednau (2003) illustrated an ovipara collected on Phyllostachys in December in China. In Taiwan it overwinters in the parthenogenetic phase (Takahashi 1923). There are conflicting reports as to whether this species does (Liao 1976), or does not (Takahashi 1921), jump when disturbed.
|
Phylloxera Boyer de Fonscolombe |
Phylloxeridae |
A genus containing nominally about 60 small pear-shaped phylloxerids living on Juglandaceae and/or Fagaceae. More than half the nominal species were described from galls on Carya in North America (Shimer 1869, as Dactylosphaera; Pergande 1904; Stoetzel & Tedders 1981). Only the pecan-feeding species have been worked on taxonomically since Pergande’s studies, and the status of many of the other Carya gall-makers is therefore uncertain. Although many species are clearly monoecious on either Carya or Fagaceae, host alternation has now been demonstrated for at least two species (Stoetzel 1985c). Accounts of the oak- and chestnut-feeding species are available for Europe (Heinze 1962, Barson & Carter 1972) and North America (Duncan 1922). Fagaceae-feeding species previously placed in the genus Moritziella Börner are now included in Phylloxera (Favret et al. 2016). Grassi (1912) fully reviewed all information on the group then available. Binazzi & Covassi (1997) compared life cycles of oak-feeding species in Italy. Hedin et al. (1985) studied the biochemistry of gall formation on pecan. Stoetzel (1985a) described the development of the sexuales, through a rapid series of moults of pupiform larvae. The nomenclatural problems associated with species described in Phylloxera and related genera are discussed by Favret et al. (2016); future work may make it necessary to formalise a distinction between most of the Palaearctic species, which have spiracles on ABD TERG 1-5, and the mainly Nearctic species, including the numerous Carya gall-makers, which only ever have them on ABD TERG 1.
Phylloxera bipunctata Lichtenstein Apterae are yellow with two orange spots, living solitarily on leaves of Quercus sp(p). in southern France and Switzerland (Lichtenstein 1874). Alatae are unknown, and the species has not been recognised since its original (brief) description.
Phylloxera caryaeavellana Riley Galls projecting underneath leaf are globular, pale greenish, like a small hazelnut (5-10 mm diam.), above leaf flat or slightly concave. Galls mature in May-June, emergent alatae have orange abdomen, BL 0.9-1.2 mm (Pergande 1904). On Carya tomentosa in Washington, D.C. The life cycle is unknown.
Phylloxera caryaecaulis (Fitch) Galls are on young twigs, on petioles or at bases of leaflet main veins, sometimes in clusters; globular, diam. 5-25 mm, pale yellowish green tinted with red before opening (see influentialpoints.com/Gallery), afterwards becoming leathery and black. Typically on Carya glabra in eastern USA. There is no host alternation. Alate sexuparae emerging from galls have dusky wings, and in life there are apparently distinctive dorsal prothoracic markings and a pair of orange or blackish spots on the posterior abdomen; BL 0.8-1.8 mm (small alatae produce mostly males, larger ones mostly females). Morgan (1909, 1912, 1915) studied the cytogenetics and chromosome cycle of this species (for summary see Blackman 1995). Shimer (1869) described three other closely-related phylloxerid species producing somewhat different twig or petiole galls on other species of Carya, which may or may not be synonymous with P. caryaecaulis: P. spinosa, producing galls covered with fleshy spines on Carya sp, in Illinois (the immature inhabitants of these galls were described by Shimer as having long spiny tubercles); P. caryaemagna, producing velvety galls on petioles or at base of mid-rib on C. amara (= cordiformis) in Illinois and Missouri; and P. subelliptica, producing ellipsoid mottled green galls on side of twig of C. alba (= ovata) in Illinois (see also Pergande 1904). Caldwell & Schuder (1979) studied the life cycle of the form on C. ovata in Indiana. Shimer (1869) reported on natural enemies of galls of P. caryaemagna in Illinois.
Phylloxera caryaefallax Riley Galls on leaves of Carya alba (= ovata) are thin-walled, 1-5 mm diam., on upper side of leaf and convex with a central nipple-like opening below leaf. Alatae with dark-veined wings and orange-yellow abdomen, BL 2.3-2.8 mm, emerge in May-early June (Pergande 1904). In Missouri, Illinois and states bordering the Mississippi at least as far south as Tenessee. The life cycle is unknown. [Morgan’s (1912,1915) cytogenetic study of Phylloxera “fallax” cannot apply to this species, as Morgan’s species was monoecious, with fundatrices that produced apterous sexuparae; its identity is still in doubt.]
Phylloxera caryaefoliae Fitch Galls on leaves of Carya glabra project above leaf as pale green slender cones with pale rings at their bases, c. 5 mm diam., almost flat on underside of leaf with a central nipple. Alatae, with distinctively marked prothorax and orange abdomen, BL 0.8-1.0 mm, emerge from the upper side of the gall (Pergande 1904) in early June. In north-eastern USA (New York, Illinois, Iowa, Virginia). The life cycle is unknown.
Phylloxera caryaeglobuli Walsh Galls according to original description are spherical, on leaf lamina between veins, opening in June to release alatae through a simple longitudinal slit on underside of leaf. On shagbark hickory (= Carya ovata) in Illinois and Missouri. Pergande (1904) recorded the available information, and suggested that P. hemispherica Shimer might be a synonym. The life cycle is unknown.
Phylloxera caryaegummosa Riley Galls are spindle-shaped, suspended from the underside of leaf of Carya alba (= ovata) by a slender petiole; they are white or yellowish, pubescent and sticky. In Illinois and Missouri, USA. Alatae are undescribed and the life cycle is unknown.
Phylloxera caryaeren Riley Galls develop in clusters along leaf petioles and mid-ribs of leaflets of Carya glabra; they are reniform, pale green, 2-15 mm max. diam., placed transversely to axis of petiole, deeply divided by a transverse slit and opening into two halves (rather like a bivalve mollusc) when mature (Pergande 1904). The phylloxerid itself is still undescribed, and the life cycle is unknown. In Illinois and Missouri, USA.
Phylloxera caryaescissa Riley Galls are globular, diam. 8-14 mm, pale greenish or dirty yellowish green, clothed with short fine yellowish hairs, projecting about eqally on both sides of leaf lamina, opening underneath when mature (mid-May to mid-June) by a transverse slit (Pergande 1904). Alatae have orange abdomen, BL 1.0-1.2 mm. On Carya tomentosa in eastern USA (Florida, D.C.). The life cycle is unknown.
Phylloxera caryaesemen Walsh Galls are rounded, small (diam. 0.3-2.0 mm) and very numerous, yellowish green to brownish yellow, rather tough and leathery, projecting more above leaf than below, with a more-or-less depressed centre above and opening (in July-early August) by a central nipple-like opening underneath. No alatae are produced; according to Walsh (1868, cited by Pergande 1904), first instar larvae emerge to found new galls. On Carya glabra in Illinois and Missouri, USA; Shimer (1869) also recorded it from C. amara (= cordiformis) in Illinois.
Phylloxera caryaesepta (Shimer) Galls of Carya alba (= ovata) have conical projections both above and below leaf, with nipple-like apices fringed with filaments when mature, the structure on both sides of the leaf somewhat resembling a miniature sea-anemone (diam. 5-12 mm). Colour of gall varies from pale yellowish green to red. Alatae with dark orange abdomen, BL 0.8-1.2 mm, emerge in late May-June from upper side of gall (Pergande 1904). In Illinois and Missouri, USA. On Carya glabra further east (New York, Virginia), the same or a closely-related species forms galls with a narrower conical structure above and below leaf (P. caryaesepta var. perforans; Pergande 1904). The life cycle is unknown.
Phylloxera caryaevenae Fitch Galls on Carya tomentosa and several other Carya spp. are distinctive, usually pale yellowish green, elongate folds along the main veins on the underside of the leaflet, with corresponding ridges or carinae above. The opening is a slit on the underside. There is no host alternation. Galls are mostly dry and empty by mid-August, but no alatae are known; the sexuales are produced from late July by apterous sexuparae in free-living colonies in deep bark fissures on the trunk and branches (Pergande 1904). Widely distributed in eastern USA, from New York to Florida and as far west as Illinois and Missouri.
Phylloxera castaneae (Haldeman) Apterae are pale yellow, BL c. 1 mm, on both sides of leaves, especially along the mid-ribs, of Castanea spp. (dentata, mollissima, pumila, sativa) in north-eastern USA (New York, Maryland, D.C., West Virginia). Infested leaves become much distorted and desiccated. Stoetzel (1985b) demonstrated host alternation in West Virginia from galls on Carya tomentosa. The galls and gall generations have not yet been described; the galls are similar to those of P. intermedia (M.B. Stoetzel, pers. comm.). Alatae from these galls gave rise to large populations on Castanea spp., causing yellow discoloration along the leaf veins. Pergande (1904) drew attention to the fact that most of the immatures on the upper sides of the chestnut leaves were bright orange (as opposed to pale yellow), and had longer tubercles than those on the undersides; his name spinifera is available for this form should it prove to be specifically distinct.
Phylloxera coccinea (von Heyden) Fundatrices in spring cause crinkling and discoloration of margins of young oak leaves. Later generations of apterae (yellowish orange, BL 0.8-1.2 mm) lay eggs in compact circles and cause necrotic spots on undersides of leaves (see Börner & Heinze 1957). Living without host alternation on Quercus spp. (especially robur) throughout most of continental Europe, and east to Kazakhstan. Both apterous and alate sexuparae are produced (Börner & Heinze 1957). Anholocyclic overwintering may occur; Lubiarz (2007) observed overwintering of immature stages of P. coccinea (and/or P. glabra) in bark crevices in Poland. P. rutila Dreyfus is a synonym (Börner 1952). New Zealand records apply to Moritziella corticalis (Sunde 1974).
Phylloxera confusa (Grassi) Fundatrices feed from veins on undersides of oak leaves, causing the lobe of the leaf distal to the feeding point to fold inwards. Later generations of apterae (BL 1.5-.2.0 mm) are yellowish to orange-yellow and develop in small groups on young twigs, on leaf scars or under loose bark (Grassi 1912). There is no host alternation; sexuparae are apterous. On Quercus robur, only recorded from central Italy.
Phylloxera castaneivora (Miyazaki) Apterae are pear-shaped, broadest anteriorly, yellowish brown to purplish brown; BL 0.9-1.3 mm. Feeding on fruits of Castanea crenata in Japan (Miyazaki 1968b, as Moritziella). Yanagibashi & Nakagaki (1985) studied the influence of temperature on development of the apterae. Other morphs and life cycle are unknown. This species was recently reported as an important pest of chestnuts in China (X. Wang et al. 2009, as Moritziella).
Phylloxera conica (Shimer) Galls are almost globular, projecting on both sides of the leaf, diameter 3-7 mm in plane of leaf, deep purplish brown, with a roughened or granular surface texture, becoming pubescent and covered with a sticky secretion when mature. Alatae, BL 0.8-1.2 mm with a pale yellow abdomen, emerge when the gall splits open underneath in June, and presumably fly to an unknown secondary host. Originally described from Carya alba (= ovata) in Illinois, USA (Shimer 1869), but the description above is of galls on C, amara (= cordiformis) in Missouri identified as this species by Pergande (1904). Riley (1875) suggested synonymy with P. depressa, but this does not accord with Pergande’s interpretation of the species.
Phylloxera corticalis Kaltenbach Apterae are pear-shaped, broadest anteriorly, green or brownish, with dark brown dorsal tubercles; BL 0.6-0.8 mm. On Quercus robur and Q. petraea, colonising bark of previous year’s growth (Heinze 1962), and sometimes causing serious dieback of shoots (Barson & Carter 1972, as Moritziella). In Europe, and introduced to New Zealand (Sunde 1973, as Moritziella). Life cycle unclear, but probably mainly anholocyclic; alate sexuparae may be produced in summer and autumn, but no sexuales have been recorded (Barson & Carter 1972). M. corticalis is barely distinct from some little-known North American species described in the genus Phylloxera (e.g. davidsoni, querceti), and may be an introduced North American species that has lost its primary host (Carya; see Stoetzel 1985).
Phylloxera davidsoni Duncan Apterae light orange to yellowish brown; BL c. 0.7 mm. Immatures are pale yellow, alatae orange with black thorax. On undersides of tomentous young leaves of Quercus engelmanni in California, USA (Duncan 1922). The life cycle is unknown.
Phylloxera deplanata Pergande Galls on leaves of Carya tomentosa are circular, diameter 1-5 mm, slightly convex above leaf and more prominent beneath, with an oval orifice fringed with fine hairs. Their colour is reddish to yellowish or greenish yellow above leaf and purplish to dull greenish yellow beneath. There is no host alternation; fundatrices produce sexuales in the galls in May, but some alatae may also be produced. Galls are empty, brown and dry by late June (Pergande 1904). Only known from D.C., USA.
Phylloxera depressa (Shimer) Galls on leaves of Carya alba (= ovata) in Illinois, USA are numerous, smooth, diameter 4-6 mm, convex and pale yellowish green above leaf, whiter beneath with a central nipple-like projection and a round opening. Alatae, BL c. 1 mm with a yellow abdomen, are present in the gall in late June (Shimer 1869). The life cycle is unknown. According to Riley (1875), conifera Shimer is a synonym.
Phylloxera devastatrix Pergande Clusters of galls occur on young twigs, leaf petioles or flower stalks of Carya illinoensis in southern and south-eastern USA (Arkansas, Louisiana, Mississippi, Oklahoma, Texas), often causing serious economic damage. The galls are globular, of irregular shape, smooth, green or yellowish green with a reddish tinge before maturity, diameter 3-15 mm. They open in May-early June by means of a transverse split into four or more broad bracts, releasing alatae that immediately start to oviposit on neighbouring twigs, etc. There is no host alternation. The eggs develop into dwarf sexuales that mate and lay overwintering eggs under bark or at the base of the tree. H. Baker (1935) described the life cycle in Louisiana, and reported a high incidence of parasitism of galls. Carpenter et al. (1979) and Calcote (1985) studied varietal and clonal resistance to P. devastatrix in pecan, Neel & Hedin (1985) studied its effects on nut production, and Hedin et al. (1985) studied biochemical aspects of gall formation. Mitchell et al. (1984) studied the gall-associated fauna.
Phylloxera foae Börner Living without host alternation on deciduous oaks, with a life cycle similar to that of Ph. coccinea, but Q. petraea is the usual host plant. Fundatrices usually in small leaf-edge folds; subsequent generations of apterae yellowish orange, BL 0.7-0.9 mm, causing necrotic spots on undersides of leaves. Sexuparae are alate. Closely related to Ph. coccinea; Heinze (1962) provided some distinguishing characters. In Italy, Germany, Ukraine.
Phylloxera foveata (Shimer) Galls on leaves of Carya amara (= cordiformis) are small (1.5-8.4 mm diam.), with a dimpled central depression above, opening beneath leaf by a rounded downy orifice. Alatae were plentiful in late June. In Illinois, USA (Shimer 1869). The same leaves carried galls of P. minima and P. caryaesemen.
Phylloxera foveola Pergande Galls on leaves of Carya glabra in Virginia, USA are circular, yellow-green tinged with pink or red, 3-5 mm diam., convex and rather prominent above leaf with a deep central dimple or depression which is pinkish in colour, part below leaf convex or depressed with a small conical central nipple. Alatae (BL c. 1.2 mm, with slender, orange abdomen) are mature in galls in late May and most had emerged before June, indicating that there is probably host alternation, but the secondary host is unknown. Possibly it is a synonym of P. foveata; the emergence time was the principal reason given for separating the two (Pergande 1904), but this might vary according to weather conditions or host plant.
Phylloxera georgiana Pergande Galls occurred singly or in clusters on tender twigs and petioles of an unidentified Carya sp., presumably at Georgiana, Florida (Pergande 1904). They were similar to those of Ph. devastatrix; more-or-less globular, with a short nipple, greenish yellow (sometimes reddened on one side), or wholly dark red, 5-10 mm in diameter. Alatae (BL 0.8-1.0 mm, with orange abdomen) differ from P. devastatrix in characters of the wings and antennae (see Pergande 1904).
Phylloxera glabra (von Heyden) Plate 2a Living without host alternation mainly on Quercus robur. Fundatrices feed in spring from main veins, sometimes causing inward curling of leaf-edge as in Ph. confusa. Subsequent generations include yellowish orange apterae (BL 0.7-0.85 mm), which lay circular clusters of eggs on the underside of the leaf, and both apterous and (later) alate sexuparae (Grassi 1912). Feeding causes necrotic spots on leaves. In Europe (England, France, Germany, Italy), and introduced to New Zealand, where it is recorded from Q. dentata (Sunde 1984). A record from Turkey (Şenol et al. 2014c) requires additional confirmation. Anholocyclic overwintering may occur; Lubiarz (2007) observed overwintering of immature stages of P. glabra (and/or P. coccinea) in bark crevices in Poland. P. glabra is regularly preyed upon by the coccinellid Pullus auritus and the coniopterygid Conwentzia psociformis.
Phylloxera globosa (Shimer) Galls on leaves of Carya amara (= cordiformis) in Illinois are mostly rather large, globular and projecting equally on both sides of leaf, or conical and projecting only below leaf, with flat circle above (“form conifera“); smooth, leathery, pale yellowish green to dark green, with a small central opening below leaf surrounded by small bracts. [According to Pergande (1904), the smaller galls included in Shimer’s (1867) original description were those of Ph. caryeaesemen.] Alatae (BL c. 0.6 mm, with orange-yellow abdomen), emerge very late in year, in late September-October. The life cycle is unknown.
Phylloxera ilicis Grassi Plate 1e, f Anholocyclic on Quercus ilex in central Italy, and alatae have apparently been trapped in Tunisia (Boukhris-Bouhachem et al. 2007). Several generations of yellow apterae occur through spring and summer feeding mainly on current and 1-year-old twigs, and causing small tuberosities; but sometimes also on foliage causing necrotic spots like P. quercus. Alate virginoparae occur in autumn generations (Grassi 1912).
Phylloxera intermedia Pergande Galls on leaves of Carya alba (= ovata) are circular, reddish or yellowish, 3-6 mm in diameter, convex to conical on upper side of leaf without any trace of a central nipple, paler and almost flat with a small closed nipple on underside. Alatae (BL 0.8-1.3 mm, with pale orange abdomen) emerge in late may-June, mainly from a jagged opening formed by splitting of the upper surface of the gall (Pergande 1904). Locality not stated; probably in vicinity of Washington, D.C. The life cycle is unknown.
Phylloxera italica Grassi Living without host alternation on Quercus robur, and only known from central Italy. Fundatrices develop in April-May and the next two generations consist only of apterae (whitish, Bl c. 1.5 mm), which feed on the undersides of leaves causing small, sparse, whitish spots. Sexuparae are mostly apterous (Grassi 1912).
Phylloxera kunugi Shinji Apterae (fundatrices?) are bright yellow, BL 2.3-2.5 mm. They feed in spring from mid-rib on undersides of young leaves of Quercus spp. (acutissima, variabilis) in Japan, causing the infested leaves to curl downward. All their progeny are alate (BL 1.8-2.5 mm), and apparently migrate to another host; the full life cycle remains to be established (Miyazaki & Teramoto 1991).
Phylloxera minima (Shimer) Galls on Carya amara (= cordiformis) in Illinois USA are small, round (diameter 1.1-5.0 mm) and numerous, with part protruding above leaf shining brightly, and a small opening beneath. Alatae, present in the galls in early July, were translucent, yellowish white with minute dark spots on anterior part of body (Shimer 1869). The life cycle is unknown.
Phylloxera notabilis Pergande Galls on leaves of Carya illinoensis are ovoid to spherical, bladder-like, very variable in size (diameter 2-18 mm), situated close to mid-rib, projecting on both sides of leaf, green on top with a central nipple, and reddish-green beneath covered with dense short yellowish hairs. Alatae (BL 0.6-0.8 mm, with dusky wings and yellow-orange abdomen), emerge from the top of the gall, which generally splits into long slender bract-like segments in June-July. There is no host alternation. Several generations of galls are produced each year; the smaller, secondary galls occurring mainly on seedling trees (Stoetzel & Tedders 1981). Brachypterous and migratory alatae, sexuparae and sexuales are all produced in the primary galls. Sexuales lay overwintering eggs on bark (Whitehead & Eastep 1937). In eastern and southern USA. Andersen & Mizell (1987) studied the physiological effects of galling on the host. Calcote (1983) studied resistance of pecan clones to P. notabilis. Medina et al. (2011) demonstrated that populations identified as P. notabilis collected from water hickory (C. aquatica)in east Texas alwayslacked two species of symbiotic bacteria that were present in populations on pecan. There were also found to be distinct differences in DNA (AFLP) profiles, and in host preferences and performance of alatae in reciprocal transfers between the two Carya species (Dickey & Medina 2010, 2011), all of which indicate that the phylloxerid on water hickory is a different, presumably undescribed, species.
Phylloxera perniciosa Pergande Galls on young twigs, leaf petioles and along midribs of leaflets of Carya tomentosa are elliptical, with a blunt tooth-like or thorn-like projection. In Maryland, Virginia, D.C. and Pennsylvania; large gall populations in Virginia caused defoliation of trees (Pergande 1904). Also reported from C. illinoensis in Louisiana and Mississippi (Carpenter et al. 1979), although the identity of these populations with Pergande’s species needs confirmation. Living all year around on Carya, with three generations per year; fundatrices, alate sexuparae and sexuales. Alate sexuparae emerging from galls in May-early June laid eggs indiscriminately on trees and surrounding vegetation. Sexuales laid overwintering eggs in June, mostly high in trees on twigs that had been previously galled. The eggs were preyed upon through the summer by various natural enemies. including mites of six different genera. Surviving eggs hatched the following April (Pergande 1904).
Phylloxera picta Pergande Galls on leaflets of Carya tomentosa resemble those of Ph. intermedia, but have thinner walls, adn a central nipple above as well as below the leaf; colour variable, diam. 3-6 mm. Alatae (BL 0.8-1.3 mm, with pale orange abdomen) emerge from an opening on the upper side in late May-June (Pergande 1904). Locality not stated; probably in vicinity of Washington, D.C. The life cycle is unknown.
Phylloxera pilosula Pergande Galls on leaflets of Carya glabra near Washington, D.C. are flat or with a central depression above leaf and projecting with a well-developed central nipple below leaf; diam. 3-6 mm, pale green to whitish, both sides densely covered with long, soft, glistening, yellowish white hairs. Alatae (BL c. 1.2 mm, with orange abdomen) emerge from the underside of the gall in early June (Pergande 1904). The life cycle is unknown.
Phylloxera querceti Pergande Apterae (= fundatrices?) are yellow to orange, BL c. 0.5 mm. On undersides of leaves of Quercus spp. (alba, dentata(?), macrocarpa) in the vicinity of Washington, D.C. (Pergande 1904), Duncan 1922). Affected leaves are speckled with small yellow spots, especially near mid-ribs and along smaller veins. Alatae of BL 0.6-1.0 mm, with orange abdomen. The life cycle is unknown.
Phylloxera quercina Ferrari (= spinulosa Targiani-Tozzetti) Apterae are pale yellow, with dorsal processes whitish (but darker in spring and autumn generations); BL 0.9-1.5 mm. On undersides of leaves of Quercus spp. (cerris, ithaburensis, suber) in the Mediterranean region (Italy, Israel, Portugal), causing yellow spots. Phylloxerids identified as this species have also been recorded from Slovenia, Turkey and Iran. Living without host alternation on Q. cerris in central Italy, 3-6 parthenogenetic generations occurring annually, with the proportion of alate sexuparae increasing in successive generations (Grassi 1912, as Acanthaphis spinulosa).
Phylloxera quercus Boyer de Fonscolombe (= florentina Targiani-Tozzetti) Apterae are greenish yellow to orange-yellow; BL 0.7-0.9 mm. On undersides of leaves of Quercus spp., causing yellowish spots. In southern Europe and south-west Asia, and heavy infestations on Q. ilex were recently reported from Algeria (Laamari 2016). (Reports from northern Europe are possibly all based on misidentification of P. glabra.) The life cycle is complex (Grassi 1912). Overwintering usually occurs as eggs on evergreen oaks, Q. coccifera or Q. ilex. Progeny of the fundatrices include alatae which fly in May to colonise deciduous oaks for the summer months (petraea, pubescens, robur), and also apterae which continue generations on Q. ilex, etc. In both cases alate sexuparae appear in the fourth and fifth generations, and those produced on deciduous oaks mostly return to evergreens to give rise to sexual morphs. Overwintering as young larvae may also occur (see Grassi 1912 and Antonelli 1989 for further details). Mohammed & Abdulla (1985) studied the ecology of populations identified as P. quercus in Iraq.
Phylloxera reticulata Duncan Apterae are bright orange-yellow, with reddish eyes; BL c. 0.8 mm. On undersides of leaves of Quercus kelloggii in California, USA, dispersed along veins. Only apterae are described (Duncan 1922), and they have not been recorded since.
Phylloxera rileyi Riley Apterae are dark brown with dorsal tubercles almost black (but paler forms mat occur); BL c. 0.6 mm. On undersides of leaves of Quercus alba and Q. obtusiloba (= stellata) in Missouri, causing white or yellow circular spots. Apparently there is no host alternation; alatae are produced from late July to October, and those of later generations are presumably sexuparae, because sexuales are recorded laying overwintering eggs on twigs (Riley 1875, Pergande 1904, Duncan 1922).
Phylloxera rimosalis Pergande Galls on Carya tomentosa in eastern USA (Washington, D.C.) are fairly large and conspicuous (diam. 3-11 mm), pale green sometimes tinged with red, usually on terminal leaves of young shoots; flat to slightly convex above leaf, more prominently convex with a central nipple beneath. Alatae (BL c. 1 mm, with pale orange abdomen) emerge in late May-June through a round opening on the underside (Pergande 1904). The life cycle is unknown.
Phylloxera russellae Stoetzel Galls on Carya illinoensis in southern USA are circular, flattened, diam. 3.2-7.2 mm, with a reticulate surface pattern. They project from both sides of a leaflet with an opening on the underside marked by white pubescence. Galls mature and break open in May. No alatae are known. There is no host alternation; apterous sexuparae produce sexuales which develop in the gall in May and produce overwintering eggs in galls, on twigs, etc. (Stoetzel 1981). Heavy populations of galls can cause defoliation. Calcote (1985) studied resistance of pecan clones to P. russellae.
Phylloxera similans Duncan Apterae of BL c. 0.7 mm, appearance in life unrecorded. Collected on Quercus macrocarpa in New York State, USA (Duncan 1922), and not recorded since the original description. Its biology is unknown.
Phylloxera spinuloides Pergande Galls on petioles or at bases of main veins of leaflets, of an undetermined Carya species in Florida, USA, were globular (diam. 5-15 mm), with a dense covering of long fleshy filamentous outgrowths; usually encircling the petiole and causing it to bend or curve. Colour uniformly greenish yellow, or brownish yellow on one side. Galls opened in late March, releasing alatae (BL 0.6-1.8 mm, with brownish wings and orange abdomen). Life cycle is unknown, although the early departure of alatae suggests host alternation.
Phylloxera stanfordiana Ferris Apterae are pale yellow, BL c. 0.75 mm. On twigs of Quercus douglasii in California, USA, in September, “concealed beneath any chance covering, especially numerous under the twig-encircling egg masses of a moth” (Ferris 1919). Other morphs and life cycle are unknown.
Phylloxera stellata Duncan Apterae (BL c. 0.5 mm, colour unrecorded) caused brown spots on undersides of leaves of a Quercus sp., margaretta (= stellata or alba, in mid-August in Texas, USA (Duncan 1922). Other morphs and life cycle are unknown.
Phylloxera symmetrica Pergande Galls on leaves of Carya tomentosa and other Carya spp. are variable; typical form is circular and almost flat above leaf, protruding as a thick-walled pale cone below leaf, basal diam. 2-5 mm. Alatae (BL 0.9-1.2 mm, with orange abdomen) emerge in late May-early June through a small opening in the apex of the cone, after which the galls become dry and brown. In vicinity of Washington D.C., USA. Life cycle is unknown. Pergande (1904) described variants from the typical form of gall (“vasculosa” and “purpurea“) differing in colour, size and shape, which he considered to be due to differences in hatching time of the fundatrices and developmental stage of the leaf at onset of feeding.
Phylloxera texana Stoetzel Galls on leaves of Carya illinoensis are globular, projecting mainly on upper side of leaf, with a small point below; diameter 4-6 mm (Stoetzel 1981). Alatae (BL 1.0-1.6 mm, with golden yellow abdomen) emerge in mid to late May through a large opening in the top of the gall, which splits into bracts. In Texas, USA. Host-alternating; alatae from galls colonised undersides of leaves of Quercus spp. (falcata, virginiana), these colonies producing both alatae and apterae in June (Stoetzel 1985b). The return migration to pecan has not been observed.
Phylloxera tuberculifera Duncan Apterae (BL c. 0.5 mm, colour unrecorded) caused brown spots on undersides of leaves of Quercus havardii in Texas, USA, in August (Duncan 1922). Other morphs and life cycle are unknown.
Phylloxera sp. Apterae are pear-shaped, dirty yellow to dark brown; BL 0.7-1.0 mm. On Quercus virginiana in Cuba, colonising apical parts of young shoots, mainly on the stem but also on young leaves which may be slightly curled or rolled. Perhaps an anholocyclic race of a North American species described in another genus (Holman 1974, as Moritziella).
|
Phylloxerina Börner |
Phylloxeridae |
About nine species mostly living without host alternation on Salicaceae, with one on Nyssa (Cornaceae). Apterae are usually covered in dense white wax. No alatae have ever been recorded for this genus. The species on Salix in Germany were studied by Iglisch (1965).
Phylloxerina capreae Börner Plate 2bApterae are yellow to brownish yellow, under dense coat of white wax filaments; BL 0.6-0.9 mm. On Salix caprea stems and branches, in bark crevices or under scales. In Europe (Germany; although a phylloxerid on Salix lanata in England, BMNH collection, leg. V.F. Eastop, has been provisionally assigned to this species), and also reported and redescribed from Japan (Miyazaki 1972b). Living without host alternation on Salix; see Iglisch (1965) for life cycle details.
Phylloxerina daphnoidis Iglisch Apterae are light brown, under dense coat of white wax filaments; BL 0.6-0.9 mm. Living without host alternation on Salix daphnoides and S. elaeagnos in Germany, in bark crevices, etc. Iglisch (1965) gave life cycle details and compared P. daphnoidis with the very similar P. capreae. The two species cannot be reliably distinguished on morphology alone.
Phylloxerina nyssae (Pergande) Galls are usually crescent-shaped, yellowish-white, formed in spring by rolling, scalloping and swelling of edges of leaves of Nyssa sylvatica (Stoetzel 1993). Later in the year (September), lemon-yellow apterae, BL 1.0-1.3 mm, completely covered in white wax wool, occur together with sexuales in bark crevices of young trees, particularly in cavities caused by dropping out of dead twigs about 2 m above ground level. Apparently there is no host alternation. In Maryland and Washington D.C., USA (Pergande 1904).
Phylloxerina popularia (Pergande) Apterae are pale greenish yellow, covered with white cottony wax, BL c. 1.4 mm, living in deserted galls of Pemphigus populitransversus and Pemphigus populicaulis, in bark crevices or on branches and twigs, of North American cottonwoods (Populus deltoides, P. fremontii and P. trichocarpa) in Louisiana, Michigan, Missouri, South Dakota, Texas and California (Pergande 1904, Davidson 1915). There is no host alternation; apterous sexuparae and sexuales were found in old Pemphigus galls in October (Pergande).
Phylloxerina populi (Del Guercio) Apterae are pale yellow to ochre, covered in wax wool; BL 0.5-0.7 mm. Alatae have not been recorded. Living without host alternation on cortical tissues of Populus spp. (alba, ×canescens). Described originally from Italy, and redescribed in detail from Hungary (Ripka 2011), who found it on branches of P. ×canescens living under scales of dead females of Chionaspis salicis (Diaspididae) along with several mite species, and also on P. alba in woody galls caused by an eriophyid mite. Bonfigli (1909; as Pseudochermes populi) and Ripka (2011) have both compared it with P. salicis.
Phylloxerina prolifera (Oestlund) Apterae are very pale lemon yellow, found in autumn in empty galls of Pemphigus populicaulis in Minnesota, USA (Oestlund 1887). Not recognised since original description, and possibly not distinct from P. popularia, despite the apparent differences noted by Pergande (1904).
Phylloxerina salicola (Pergande) Apterae are yellow to brownish yellow, covered with white wax wool; BL 0.6-0.9 mm. Originally described from bark crevices or other cracks in stems and twigs, and sometimes also on undersides of leaves, of an unidentified Salix species (either discolor or humilis) in eastern USA (Pergande 1904). Sexuales are present in September. Davidson (1915) recorded this species from Salix lasiolepis in California, usually living in bud axils on twigs but also on one occasion heavily infesting bared roots on a stream bank. These Californian populations appeared to be anholocyclic, and could possibly have been introduced European P. salicis.
Phylloxerina salicis (Lichtenstein) Apterae are pale green, yellow or brownish, secreting profuse white wax wool; BL 0.7-1.2 mm. Living in bark crevices on trunk and branches of Salix alba and related species in western, southern and central Europe, and across temperate Asia to China (G. Zhang et al. 1985) and east Siberia (Pashtshenko 1988a). Introduced to Victoria, Australia (Eastop 1966), and perhaps to North America (Börner & Heinze 1957). Sexuales occur in September (Iglisch 1965).
Phylloxerina sp. Holman (1964) described (but did not name) a species of Phylloxerina on trunks of Salix longipes in Cuba. The life cycle is unknown. Similar to European P. capreae (but not compared with the North American species).
|
Pineus Shimer |
Adelgidae |
Clearly differentiated from the other adelgid group, Adelges, by their biology – e.g. gall structure, secondary host association with Pinus – and by various morphological features. The most consistent character that applies to all morphs is the reduction in the number of abdominal spiracles to only four evident pairs. The galls, usually on shoot tips of Picea, are generally less compact than those of Adelges and have intercommunicating chambers. Adult apterous morphs on secondary hosts (sistens and progrediens) are very broadly pear-shaped – often almost globular – and have a fused and pigmented head and prothoracic shield.
Pineus species may be holocyclic and host-alternating between Picea and Pinus, completing a cycle with seven morphs in two years as in holocyclic Adelges; or they may be anholocyclic on either Picea or Pinus (except for Pineus abietinus, which is anholocyclic on Abies). There is also one species, Pineus floccus, that is now known to have an anholocyclic, annual host alternation between Picea and Pinus. Knowledge of the life cycle of many species is still incomplete. Several anholocyclic species on Pinus produce alate sexuparae which appear to be non-functional, either because they are unable to produce males or because they fail to find a suitable Picea species to act as primary host.
Systematic accounts of Pineus are available for Britain (Carter 1971), Europe (Börner & Heinze 1957, Heinze 1962), India (A.K. Ghosh 1984a), Japan (Inouye 1953) and North America (Annand 1928). Favret et al. (2015b) provided a taxonomic and nomenclatural catalogue of the world fauna. Havill et al. (2007) studied the evolution of host relationships based on molecular data. Havelka et al. (2019) analysed mitochondrian and nuclear DNA sequnces of the species in Lithuania.
Pineus abietinus Underwood & Balch (fig. 10A) Only the first instar and adult progrediens (aestivoprogrediens) are described; adults are very small (about 0.6 mm), light reddish-brown with dark cephaloprothoracic shield, secreting white wax-wool, in bark crevices on stems and branches of Abies amabilis and A. grandis in the north-west coastal region of North America (Washington, southern British Columbia). This is the only known Pineus associated with Abies. Wax pores are circular and discontiguous. Apparently anholocyclic and monomorphic, probably with two or three progrediens generations per year (Underwood & Balch 1964).
Pineus armandicola Zhang, Zhong & Zhang Galls on Picea likiangensis in Yunnan, China are cone-like or torch-like with multiple cavities (Li et al. 2002). Migration occurs to Pinus armandii.Apterae on Pinus are red-brown, with wax threads; BL 0.8-1.4 mm (G. Zhang et al. 1992c).
Pineus boerneri Annand Apterae are small, reddish brown, laying eggs on pine twigs in abundant greyish-white wax wool. Described from Pinus radiata in California (Annand 1928), but possibly of east Asian origin, where it has mainly been recorded under the name P. laevis (McClure 1984b). It is now also in north-eastern USA, Hawaii, South America (originally as P. havrylenkoi Blanchard), Australia, New Zealand and east Africa (as P. pini). P. simmondsi Yaseen & Ghani, on Pinus roxburghii in Pakistan (Yaseen & Ghani 1971), could also be this species. P. boerneri is anholocyclic everywhere, and the introduced populations attack a range of exotic pines from both Europe (e.g. P. halepensis) and North America (e.g. P. radiata, P. caribaea). In Australia and New Zealand the name laevis has been applied to both boerneri and pini (q.v.), which are difficult to differentiate except by using multivariate morphometrics and karyotype analysis (Blackman et al. 1995). Biological control of boerneri and/or pini has been successful using Leucopis sp. (Chamaemyidae) in New Zealand, Chile and Hawaii (Mills 1990). Dynamics of east African populations was studied by Mailu et al. (1980, as pini), and in the rather different conditions of Pinus resinosa plantations in Connecticut, USA by McClure (1989b, 1990). Cardoso (2007) has made a detailed study of its morphology, ecology and feeding behaviour in Brazil. 2n=16 (Hawaii) or 17 (California, Africa, Australia, New Zealand).
Pineus boycei Annand (fig. 64B) This species forms 2-4 cm long galls on Picea engelmanni, green with a reddish tint, at the bases of the needles, usually at the shoot tips. It is only recorded from Montana and Oregon, USA. Alate gallicolae emerging from the galls in the second half of July are 1.7-2.5 mm, with abundant wax posteriorly on the reddish brown abdomen (Annand 1928). Their destination is unknown. No other morphs of life cycle details are known (although it seems possible that alate specimens described by Annand under the name Pineus engelmanni are the sexuparae of this species).
Pineus cembrae (Cholodkovsky) (= sibiricus Cholodkovsky) Host-alternating between Picea spp. (abies, glehnii, obovata) and stone pines, Pinus spp. of subsection Cembrae (cembra, pumila, koraiensis, sibirica); of boreo-alpine distribution in Europe, and extending across Asia to Japan. Other pines are not usually attacked (Potopova 1984), except that P. parviflora is recorded as a host in Japan. Galls are 4-5 cm long, often asymmetrical, distorting shoot tips, with needles concentrated on one side. Gallicolae are dark red and secrete abundant wax, migrating to Pinus in June in Germany (Börner & Heinze 1957), but in August in Japan (Inouye 1953). The neosistens overwinter mainly near ends of twigs at the bases of last year’s pine needles, giving rise the following year to alate sexuparae and one or two generations of apterous progredientes living in dense wax-wool on new shoots. Inouye (1953) describes and illustrates all morphs. A form collected on the needles of P. koraiensis in Heilongjiang Province, China is described as a subspecies, P. cembrae pinikoreanus (G. Zhang & Fang 1981). 2n=18.
Pineus cladogenous Fang & Sun Anholocyclic on Pinus koraiensis in Heilongjiang, China, overwintering in fissures in the bark. The adult hiemosistentes (collected in April) are small and ladybird-shaped; BL about 0.6 mm (Fang & Sun 1985). Perhaps closely related to P. harukawai.
Pineus coloradensis (Gillette) Anholocyclic on numerous Pinus spp. in USA and Canada. Apterous progredientes feeding on the needles secrete abundant wax, which in summer becomes covered with a dark mould. One hiemoprogrediens and two aestivoprogrediens generations occur in New England, where this adelgid has been injurious to plantations of P.resinosa (McClure 1989b). Alate sexuparae, presumed non-functional, are recorded from Colorado, but not from California or Connecticut. Descriptions of morphology and biology in comparison with related species are given by Annand (1928), Doane (1961) and McClure (1989b). McClure (1984a) studied interactions between this adelgid and Pineus boerneri on Pinus resinosa, and Huff & McDonald (1977) studied differences in its susceptibility to numerous white pine species.
Pineus cortecicola Fang & Sun Anholocyclic on Pinus koraiensis in Heilongjiang, China, overwintering under bark in only a little wax (Fang & Sun 1985). BL of adult hiemosistens about 0.85 mm. From the description this species appears similar to P. hosayai, but differs in overwintering site and amount of wax.
Pineus floccus (Patch) “Host-alternating” between Picea spp. (rubens, mariana) and Pinus strobus in eastern USA, but the host alternation revealed by life history studies in Virginia is remarkable because it occurs on an annual basis, and only involves parthenogenetic morphs; there are no fundatrices, sexuparae or sexuales (Walton 1980). There is a single generation (gallicolae) on Picea, and a series of progrediens generations on pine. Overwintering apterous progredientes on P. strobus produce a spring generation of both apterous and alate progredientes, and the latter fly to spruce buds in May. Galls are induced by the developing gallicolae. They are usually terminal, 1.8-4.5 cm long, resembling those of Adelges cooleyi but shorter, stouter and looser; green, but acquiring a purplish or reddish tinge as they mature, the protruding ends of the needles remaining green and of normal length. Adult gallicolae (fig. 64D), 0.9-2.0 mm with abundant wax on head and thorax and a yellow-brown abdomen, migrate in July to both old and new needles of Pinus strobus. Apterous adults of the second generation on pine overwinter at the bases of buds, needles or stem scales, moving to growing points in spring. In New York, however, where heavy galling of Picea rubens in plantations is recorded, Deboo & Weidhaas (1967) believed that gallicolae were recolonising spruce directly and not migrating to pine.
Pineus ghanii Yaseen & Ghani Anholocyclic on Pinus griffithii in the Galis and Kaghan valleys, Pakistan, attacking stems and twigs of mature and old trees (Yaseen & Ghani 1971). There are two generations per year, a hiemosistens becoming adult in April, and an aestivosistens becoming adult in August-September. Adults are light reddish-brown, 0.9-1.2 mm.
Pineus harukawai Inouye Anholocyclic on white pines (Pinus parviflora, P. strobus) in Japan (Hokkaido, Honshu, Kyushu), on stems or older branches of young pines, or at the bases of the needles on young branches of older pines. Only the adult of the overwintering sistens – about 0.9 mm and orange to brownish-red with abundant wax, laying large numbers of eggs in April to May – is described; other details of the life cycle are unknown. This species is sometimes injurious in pine nurseries in Hokkaido (Inouye 1953).
Pineus hosoyai Inouye Only the apterous rogrediens and immature sexupara are described, collected from Pinus koraiensis in June in Korea, and from Pinus sp. in May in China (Inouye 1953). The overwintering sistentes were found in February in Korea, living in abundant wax on shoots of the previous year’s growth. The life cycle is unknown.
Pineus matsumurai Inouye Apterous progredientes and alate sexuparae develop in June at needle bases on young branches of Pinus thunbergii and P. densiflora in Japan (Inouye 1953). The apterous progrediens is small (about 0.8 mm), dark reddish-brown, covered in dense fine wax-wool. The destination of the sexuparae and the rest of the life cycle is unknown. This species seems to be close to Pineus strobi; differences are discussed by Inouye (1953).
Pineus orientalis (Dreyfus) (fig. 64C) Host-alternating between certain Picea spp. (orientalis, polita, jezoensis, but not abies) and various Pinus spp. of the subsection (densiflora, mugo, sylvestris, thunbergii) from western Europe across Asia to Japan. Galls are terminal, shorter and usually straighter than those of P. cembrae, with protruding needle tips evenly distributed around the gall. Gallicolae (Plate 1d, fig. 64E) fly to Pinus in late June to early July and lay egg-masses in wax wool on the previous year’s needles. There are numerous acounts of the biology of this adelgid, which is not differentiated from its anholocyclic close relative P. pini in some of the earlier literature (e.g. in Europe: Börner 1908, Marchal 1913, van der Goot 1915, Mordvilko 1935; and in Japan: Inouye 1953). 2n=20.
Pineus pineoides (Cholodkovsky) (fig. 64A) Anholocyclic, with one sumer and one winter generation per year, both of apterae which live on well-shaded parts of the bark of the trunk and larger branches of Picea spp. (abies, obovata, omorika, rubens). The insects are yellowish-grey to dark red-brown, dorso-ventrally flattened, with the cephaloprothoracic shield of strongly pigmented specimens heavily sclerotised; BL 0.6-0.8 mm. Each lays its eggs under a separate tuft of wax wool. The overwintering generation becomes adult in May. The adults of the summer generation, similar except that they have rather fewer wax pores, may be found in July. In Europe, Turkey (Şenol et al. 2014), Canada, and also recorded from bonsai spruce in Australia (New South Wales; BMNH collection, leg. D. Koreshoff). For details of biology see Steffan (1963, in Germany) and Underwood (1963, in Canada). Pineus konowashiyai, on Picea abies in Japan, has a similar habitat and life cycle to P. pineoides, but differs in having large discontiguous wax pores (Inouye 1953). 2n=22 (Steffan, 1968) or 17 (Blackman & Eastop 1994).
Pineus pini (Macquart) Plate 1c, fig. 72BAnholocyclic on Pinus sylvestris and Pinus mugo in western, northern and central Europe, with an overwintering (hiemosistens) generation on the twigs and two or more overlapping generations of progredientes attacking the current year’s shoots. Apterae are dark brown or dark red, BL 1.0-1.2 mm, secreting copious wax-wool, in which the eggs are laid (see influentialpoints.com/Gallery). The second progrediens generation in May-June includes alatae, but first instar crawlers dispersed by wind may be a more common means of infection of young seedling pines (C.I. Carter, pers. comm.).
The morphs of P. pini are all virtually indistiguishable from the equivalent secondary host generations of P. orientalis, and it seems probable that the anholocyclic P. pini arose from the holocyclic P. orientalis by loss of a functional sexual generation, as suggested by Marchal (1913), and supported by mitochondrial and nuclear DNA sequence analysis (Havelka et al. 2019). P. pini in Europe seems to restrict its attacks mainly to Pinus sylvestris and P. mugo, but anholocyclic populations introduced to the southern hemisphere can be injurious to a wide range of pine species. In the Australian literature populations of P. pini group have commonly been given the name laevis Maskell and confused with P. boerneri, which also occurs on a range of pines in both Australia and New Zealand. P. pini has also been recorded from pine plantations in east Africa but these records should all be referred to P. boerneri (Blackman et al. 1995). 2n=20, 21 (Blackman et al. 1995) or 2n=22 (Steffan 1968).
Pineus pinifoliae (Fitch) (fig. 72A) Host-alternating between Picea spp. (engelmanni, glauca, mariana, pungens, rubens, sitchensis) and white pines (Pinus strobus, P. monticola) in North America. The galls on spruce are conical, 1.8-4.3 cm long, with their constituent needles characteristically flattened and broadened to form “scales”. They are purple and green at first, becoming reddish-brown at the time of opening in mid-june to early July, when the needle-scales flare outwards. The gallicolae, 1.6-2.3 mm, fly to needles of the previous year’s growth of white pine; they invariably face the base of the needles and deposit their eggs beneath their wings, dying after oviposition with the wings still protecting the eggs. The first instar sistentes (neosistentes) move to the axes of new shoots to overwinter. There are two apterous progrediens generations on pine, alate sexuparae being produced in May to fly to old needles of spruce. For further life cycle details see Balch & Underwood (1950). Dynamics of the gall populations on spruce was studied by Howse & Dimond (1965), and of the populations on white pine by Dimond & Allen (1974). The histology of feeding damage to white pine shoots has also been studied (Allen & Dimond 1968). 2n=22 according to Steffan (1968).
Pineus piniyunnanensis Zhang, Zhong & Zhang Apterae red with black head and prothorax, presumably secreting wax in life; BL c. 0.8 mm. On Pinus yunnanensis in Yunnan, China (G. Zhang et al. 1992c). The life cycle is unknown.
Pineus sichuananus Zhang Described from Picea likiangensis and Picea purpurea in Sichuan, China. The first instar and adult sistens, which one would normally expect to occur on a Pinus species, were recorded along with galls and gallicolae on spruce, so the life cycle needs clarification. P. sichuananus is considered to be closely related to P. cembrae; the differences given all involve the wax glands (G. Zhang et al. 1980).
Pineus similis (Gillette) Anholocyclic, living without host alternation on a wide range of Picea spp. in North America (abies, glauca, engelmanni, mariana, pungens, rubens), and introduced into Britain where it also colonises Picea sitchensis (Carter 1975). The unusual life cycle of this adelgid was investigated in Canada by Cumming (1962a). Pseudofundatrices (not true fundatrices, because they hatch from unfertilised eggs) overwinter in the second or third instar under bud scales. Their feeding in spring causes rather loose galls 1-4 cm long, with the constituent needles much paler than the normal new growth (see bladmineerders.nl/gallen). As the gall matures the shoot and needles become brown and twisted giving it a ragged appearance. Uniquely, both alate and apterous gallicolae are produced in the galls. Apterous gallicolae oviposit within the galls, but apterae may also be found outside the galls, laying eggs in small tufts of wax on the stem. Alate gallicolae, 1.0-2.1 mm and dark reddish-brown, fly in June-July to recolonise spruce needles, laying eggs in white wax wool. These apparently develop into the overwintering pseudofundatrix generation. 2n=22 according to Steffan (1968).
Pineus strobi (Hartig) Anholocyclic on Pinus strobus where this pine species is native in eastern North America, and introduced on this pine into Europe. In England, Carter (1971) has recorded it attacking one species of European white pine, Pinus peuce, and in the BMNH collection there are also specimens tentatively identified as this species from P. cembra, P. pinea and P. sylvestris. It has also now been recorded from western Siberia (Altai Republic) on Pinus sibirica (Stekolshchikov & Novgorodova 2013). Recently it is also reported from western North America (Darr et al. 2018). Raske & Hodson (1964) describe the life cycle and biology in Minnesota, USA, and their observations largely agree with those made in Europe (e.g. Marchal 1913, Carter 1971). Overlapping generations of sistentes and progredientes feed through the spring and summer on shaded parts of the bark of the trunk and undersides of branches, secreting copious white wax wool. On seedling trees, parts of the new growth or last year’s growth wher the needles are densest are colonised. Overwintering occurs as immaure apterae (progrediens or sistens) on the trunk, or under scales of dwarf-shoots. Alate sexuparae are often produced in May-June, and fly to Picea mariana to lay eggs on the needles. However, the sexual phase is abortive, either because the sexuales hatch but fail to develop (Raske & Hodson 1964) or due to the absence of any males (Marchal 1913). No galls or gallicolae of P. strobi have been found on Picea mariana or any other spruce. Sloss & Foote (1973) studied predation by Leucopis pinicola (Chamaemyiidae) and other natural enemies of P. strobi in Ohio, USA, and Wantuch et al. (2019) investigated its predator complex in Virginia, USA. 2n=22 (Steffan 1968) or 20 (Blackman & Eastop 1994).
Pineus sylvestris Annand Apparently anholocyclic in Pinus spp. (radiata, sylvestris) in California, USA (Annand 1928). Inouye (1953) records what appears to be the same species from Pinus thunbergii in Hokkaido, Japan. Reddish-brown apterous progredientes, 1.0-1.1 mm with rather sparse wax, were found in May-June in California feeding inconspicuously at the needle bases near male cones. Alate sexuparae appeared early in May, but their fate is unknown, no generations on spruce having been identified. The overwintering stages on Pinus, mainly second and third instar progredientes, were found under stem scales and behind lateral buds near the ends of branches.
Pineus wallichiana Yaseen & Ghani Anholocyclic on Pinus griffithii (= wallichiana) in the Galis and Swat Hills, Pakistan, attacking stems and twigs of mature and old trees (Yaseen & Ghani 1971). Adult sistentes are about 0.9-1.0 mm, light brown with a thick covering of wax. There are only one or two sistens generations per year, and no alatae are known. Morphology and life cycle are apparently very like Pineus ghanii, and this is possibly a highland form of that species.
|
Platyaphis Takahashi |
Calaphidinae: Calaphidini (?) |
One species on Fagus in Japan. Only the very specialized apterous vivipara is known, and its phylogenetic position is uncertain. Takahashi (1957) compared it with Dasyaphis, but Shaposhnikov & Gabrid (1987) suggested that it belongs in the Hormaphidinae.
Platyaphis fagi Takahashi (fig. 47A) Apterae are broadly ovate, flattened, green with pale antennae and legs; BL 1.8-2.0 mm. On undersides of leaves, along mid-ribs, on Fagus crenata in Japan (Takahashi 1957). Life cycle and other morphs are unknown.
|
Pleotrichophorus Börner |
Aphidinae: Macrosiphini |
About 60 species of mainly pale green to yellow aphids bearing numerous short capitate hairs, and with slender, often elongate appendages. With only a few exceptions the species live on Compositae, mostly on Anthemideae (29 spp., of which 24 are on Artemisia and 4 on Achillea) and Astereae (19 species, of which 15 are on Chrysothamnus or Ericameria), with a few species on Inuleae (Gnaphalium, Helichrysum). They are specialised feeders on plants with dense glandular hairs. The genus is predominantly American (49 species), with seven European and only three central Asiatic and one east Asian species known. Many of the species were originally described in Capitophorus. Daniyarova & Narzikulov (1984) reviewed the genus, and regional accounts are available for Europe (Holman 1965a, Leclant 1968a, Heie 1994, Blackman 2010) and America (Hille Ris Lambers 1969, Corpuz Raros & Cook 1974). Kadyrbekov (2016) provided a key to the palaearctic species.
Pleotrichophorus achilleae Holman Apterae are greenish or pinkish white, with dusky to dark appendages; BL 1.6-2.3 mm. On undersides of leaves of Achillea pectinata (= kitaibeliana) in eastern Europe (Slovakia, Hungary). Monoecious holocyclic with oviparae and apterous males in October (Holman 1965a).
Pleotrichophorus ambrosiae Hille Ris Lambers (Fig.8a) Apterae are pale green to whitish green with dusky apices to antennae and legs, BL 1.8-2.3 mm. On undersides of leaves of Ambrosia spp., widely distributed in USA , and in Mexico and Cuba (Holman 1974b). Alatae have no central dorsal abdominal patch (cf. Capitophorus spp.). Monoecious holocyclic in USA (e.g. Colorado), with alate males (Corpuz-Raros & Cook 1974).
Pleotrichophorus amsinckii Richards Apterae are pale green, appearing “weakly pulverulent” due to dense dorsal hairs; BL 1.6-1.8 mm. Described from Amsinckia intermedia in British Columbia, Canada and subsequently recorded from Washington, USA on Eriogonum sp., Phacelia hastata and Eriophyllum lanatum, which makes the true host relationship uncertain.
Pleotrichophorus antennarius Corpuz-Raros & Cook Colour of apterae in life is unknown; BL 1.1-1.6 mm. On Antennaria plantaginifolia and Antennaria sp. in Washington D.C., USA. Monoecious holocyclic with oviparae and apterous males in October (Corpuz-Raros & Cook 1974).
Pleotrichophorus artemisicola (Williams) Apterae are pale green with pulverulent appearance due to dense, capitate hairs, with mainly dark antennae and legs, black siphunculi and white cauda; BL 1.7-1.9 mm. On upper part of stem and leaves of Artemisia cana. This specie is noted to be very active. Described from Nebraska, USA, and there are also records from California, Oregon and Kansas.
Pleotrichophorus asterifoliae (Strom) Apterae are shiny dirty white to pale yellow with antennae and legs distally dark; BL 1.8-2.4 mm. On Aster sp. in northern USA and Canada, forming large colonies on undersides of lower leaves of plants growing in shady areas. Oviparae were found in autumn in Wisconsin, USA (original description).
Pleotrichophorus blackmani Barjadze, Halbert & Kanturski Apterae are light green; BL 1.5-2.1 mm. On Euthamia graminifolia in Florida (USA) and perhaps in Ontario (Canada). Oviparae occur in January (original description).
Pleotrichophorus brevinectarius (Gillette & Palmer) Apterae are mid-green, with frosted appearance due to dorsal hairs; BL 1.3-2.0 mm. On leaves and stems of Artemisia longifolia, and and subsequently recorded from several other Artemisia spp. In western USA.
Pleotrichophorus chrysanthemi (Theobald) Plate 16f Apterae are pale green to yellowish (see aphids of Karnataka website and influentialpoints.com/Gallery); BL 1.8-2.4 mm. On undersides of leaves of florists’ chrysanthemums, (Dendranthema spp.), often forming large colonies on yellowing leaves. Described from South Africa, and since recorded from Europe, Egypt, Zimbabwe, Iran, India, Nepal, Korea, Japan, Australia, North and South America. Capitophorus formosanus Takahashi, recorded from Japan and Taiwan on Chrysanthemum, is possibly this species. Apparently it is entirely anholocyclic; no sexual morphs are known. [P. chrysanthemi is difficult to distinguish from P. glandulosus and may not deserve separate species status. Alatae from Dendranthema generally have fewer secondary rhinaria (9-20 on ANT III and none on IV) than those from Artemisia vulgaris, which have 14-23 on ANT III and 0-8 on IV (most with 3 or more on IV). Best discriminant for apterae is perhaps the number of hairs on abdominal tergite 8; P. chrysanthemi has 5-11 (64% with 7-8), and P. glandulosus has 8-13 (65% with 9-10). Populations of this species group on Artemisia apiacea in China (BMNH collection, leg. V.F. Eastop) resemble chrysanthemi rather than glandulosus, and Lee et al. (2002c) also identify populations on Artemisia princeps var. orientalis in Korea as chrysanthemi; it seems likelythat chrysanthemi may have arisen quite recently in east Asia as an anholocyclic form or clone able to feed on Dendrathema, and subsequently dispersed widely on florists’ chrysanthemums.]
Pleotrichophorus decampus (Knowlton & Smith) (Fig.14k) Apterae are bluish green to apple-green with “whitish cast” due to dorsal hairs; BL 1.3-2.0 mm. On Artemisia californica and A. tridentata in western USA and Mexico. Records from other Artemisia spp. are mostly assignable to P. pseudoglandulosus, with whichthere is much confusion in the literature. Oviparae and alate males of P. decampus occur on A. tridentata in Utah in October (Corpuz-Raros & Cook 1974).
Pleotrichophorus deviatus Müller Apterae are dull whitish; BL 1.8-2.4 mm. On undersides of leaves of Hieracium spp. in continental Europe. Oviparae and apterous males occur in September (original description).
Pleotrichophorus diutius Corpuz-Raros & Cook Apterae are green, BL 1.6-2.2mm. On leaves of Artemisia dracunculoides and Artemisia sp. in USA (New Mexico, Colorado, Oregon, Minnesota). This species is confused with P. wasatchii in Palmer (1952); see Corpuz-Raros & Cook (1974).
Pleotrichophorus duponti Hille Ris Lambers Apterae are dull greyish greenwith green transverse stripes; BL 1.6-2.1 mm. On lower leaves of Achillea spp., falling when disturbed. In western, northern, central and eastern Europe. Ossiannilsson (1959) recorded it from Leucanthemum vulgare.Monoecious holocyclic on Achillea with oviparae and apterous males in September-October (Hille Ris Lambers 1953). 2n=14.
Pleotrichophorus elongatus (Knowlton) (Fig.21f) Apterae are shining apple green, slenderly spindle-shaped; BL 1.7-2.9 mm. On leaves and apical parts of Chrysothamnus and Ericameria spp. in western USA. As suggested by Palmer (1952) and Hille Ris Lambers (1969), P. palmerae (Knowlton) is probably a synonym; the characters given to distinguish it from elongatus by Knowlton, and by Corpuz-Raros & Cook (1974), are those likely to be found in alatiform specimens.
Pleotrichophorus filaginis (Schouteden) Apterae are purplish grey, BL 1.5-2.2 mm. Alatae have dark brown abdominal pleural intersegmental sclerites. On Filago and Gnaphalium spp., and also recorded from Helichrysum arenarium. They feed solitarily or in small single-parent families on upper- and undersides of the leaves. Monoecious holocyclic, producing oviparae and apterous males in October (Hille Ris Lambers 1953).
Pleotrichophorus filifoliae (Palmer) Apterae are small, mid-green , with frosted appearance due to dorsal hairs; BL 1.0-1.3 mm. On leaves of Artemisia filifolia in Colorado, USA. Recently it has been collected from Artemisia sp(p). in New Mexico, Utah, Idaho and Oregon, including oviparae found on A. tripartita in October in Oregon (A. Jensen, aphidtrek.org).
Pleotrichophorus glandulosus (Kaltenbach) (Fig.14e) Apterae are yellowish white, sometimes greenish, occasionally with a pale green median stripe; BL 1.4-2.6 mm. On undersides of lower leaves of Artemisia vulgaris, and sometimes on other Artemisia spp. Records from other genera are probably of vagrants, or based on misidentifications. In Europe and across Asia to Japan, China, Korea and Taiwan, and introduced to eastern North America; records from western North America are possibly all due to confusion with other species (e.g. decampus, gnaphalodes, pseudoglandulosus). Also in Argentina (Ortego et al. 2004) and Uruguay (BMNH collection, leg. V.F. Eastop); but alatae from A. verlotiorum in Uruguay resemble P. chrysanthemi (q.v.) in lacking secondary rhinaria on ANT IV. Oviparae and apterous males of P. glandulosus occur on Artemisia vulgaris in September-October (Heie 1994, Corpuz-Raros & Cook 1974). Nevsky’s (1929) subspecies in Central Asia, P. glandulosus ssp. ispharinus, is probably just a variant population. However another variant population with fundatrix, apterae and oviparae all found in July on Tanacetum bipinnatum in north-west Russia (Nenets region) and described as P. sp. aff. glandulosus by Stekolshchikov (2017) is possibly a distinct species. 2n=14.
Pleotrichophorus gnaphalodes (Palmer) (Fig.14f,h) Apterae are pale green with frosted appearance (see influentialpoints.com/Gallery); BL 1.3-2.2 mm. On undersides of leaves of Artemisia douglasiana, A. dracunculus, A. ludoviciana and A. tridentata in western USA, and in Mexico on A. mexicana (BMNH collection, leg. G. Remaudière).
Pleotrichophorus gregarius (Knowlton) (Fig.21b,e) Apterae are brown to greenish brown in summer and autumn, with antennae black at apices of III and IV and dark on V-VI, and siphunculi pale at bases but dark distally; BL 1.8-3.2 mm. Spring generations are dark brown to chocolate brown. On growing points and leaves of Chrysothamnus and Ericameria spp., with especially large, dense colonies developing on rabbit brush (E. nauseosa), in western USA. Oviparae and alate males occur in October-November (original description).
Pleotrichophorus helichrysi Bozhko (Fig.31 l,n) Apterae are transparent white or pale greenish, with dull greyish appearance due to the numerous fan-shaped dorsal hairs; BL 1.5-2.1 mm. Living well-camouflaged on undersides of leaves of Helichrysum arenarium.In Denmark, Poland, Czech Republic, southern Russia and Kazakhstan (Bozhko 1976b, Heie 1994, Kadyrbekov 2017a).
Pleotrichophorus heterohirsutus (Gillette & Palmer) (Fig.14g) Apterae are pale green with frosted appearance due to dorsal hairs; BL 1.0-1.8 mm. On leaves and stems of Artemisia tridentata, forming crowded colonies on young plants, in western USA (Colorado, Idaho, Utah, Wyoming). Oviparae and apterous males occur in September (Palmer 1952).
Pleotrichophorus hottesi Hille Ris Lambers Apterae are greyish green; BL 1.9-2.3 mm. On Achillea lanulosa (= millefolium?)in North America (Colorado, North Carolina, Manitoba). Monoecious holocyclic with apterous males in September (original description).
Pleotrichophorus infrequenus (Knowlton & Smith) (Fig.14a) Apterae are pale (colour unrecorded), BL c.1.3-1.4 mm. On leaves of Artemisia tridentata in western USA (Idaho, Utah, Oregon) and Mexico (BMNH collection, leg. G. Remaudière). Specimens collected from Artemisia rigida (Oregon: BMNH collection) have a rather larger cauda, and may represent a different taxon.
Pleotrichophorus knowltoni Corpuz-Raros & Cook Colour in life is unknown, probably pale with frosted appearance; BL c. 2.5 mm. Described from an aptera and oviparae collected in July on an unidentified host in Alaska, and subsequently found on Artemisia spp. including A. tilesii in north-east Siberia (Chukotka) by Stekolshchikov & Khruleva (2015), who provided detailed descriptions of fundatrices and apterous males collected in July-August. This species is very similar to P. glandulosus, but with more numerous dorsal hairs.
Pleotrichophorus lagacei Hille Ris Lambers Colour of apterae in life is unknown, probably very pale with frosted appearance; BL 1.8-2.5 mm. On Ericameria bloomeri at an altitude of 2,600m in California, and also recorded from Oregon (A. Jensen, aphidtrek.org).
Pleotrichophorus longinectarius (Gillette & Palmer) Apterae are pea-green, with frosted appearance; BL 2.0-2.5 mm. On leaves and stems of Artemisia longifolia and Artemisia sp. in western USA (Utah, Wyoming, New Mexico) and Canada (Alberta, Manitoba).
Pleotrichophorus longipes (Gillette & Palmer) (Fig.14b) Apterae are bluish apple green, and apparently wax-dusted; BL 2.4-3.0 mm. Living singly and well-camouflaged on leaves of Artemisia tridentata in western USA (California, Colorado, Idaho, Utah, Wyoming). Oviparae and apterous males occur in September (Palmer 1952).
Pleotrichophorus longirostris Hille Ris Lambers Colour in life is unknown; BL 1.6-1.9 mm. On Eriophyllum stoechadifolium in California, USA, and also on Eriophyllum sp(p). in Washington and Oregon (A. Jensen, aphidtrek.org). This species has fan-shaped dorsal hairs and a long stiletto-shaped R IV+V (0.17-0.19 mm), similar in length to the short thin siphunculi.
Pleotrichophorus magnautensus (Knowlton & Smith) (Fig.21c) Apterae are green, with short black siphunculi; BL 2.3-2.7 mm. On Chrysothamnus viscidiflorus in Utah, Wyoming and Colorado, USA. Alatae are undescribed.
Pleotrichophorus narzikulovi Umarov Apterae are green, apparently dusted with grey wax; BL 2.5-3.0 mm. Living in small colonies on Artemisia dracunculus in Central Asia (Kyrgyzstan). Alatae are unknown.
Pleotrichophorus obscuratus Hille Ris Lambers Apterae are greyish green, apparently waxy due to dorsal hairs, with dark antennae, legs, apices of siphunculi, cauda and anal plate, and sometimes conspicuously dark subgenital plate; BL 1.3-1.8 mm. Described from Artemisia californica in California, and subsequently collected on several other Artemisia spp. in Oregon, Utah, Nevada and New Mexico (A. Jensen, aphidtrek.org). Alatae are undescribed.
Pleotrichophorus oestlundi (Knowlton) (Fig.21h) Apterae are bluish green to apple green, with apparent white pruinosity due to dense dorsal covering of fan-shaped hairs (see influentialpoints.com/Gallery); BL 1.5-2.3 mm. On Chrysothamnus and Ericameria spp. in western USA. Oviparae (red to brownish red in life) and alate males occur in October (original description).
Pleotrichophorus ohioensis Smith Apterae are greenish with greyish white overcast due to fan-shaped hairs; BL 2.1-2.4 mm. On undersides of leaves of Helianthus sp. in Ohio, USA. Oviparae and alate males were collected in mid-October (original description).
Pleotrichophorus packi (Knowlton) (Fig.21g) Apterae are rather slenderly spindle-shaped, bluish green to apple green or yellow, appearing rather shiny or frosted due to fan-shaped dorsal hairs, with long dark siphunculi, curved outward apically; BL 1.9-3.5 mm. On leaves of Chrysothamnus and Ericameria spp. in western USA. Oviparae and alate males occur in late October-November (Palmer 1952). Corpuz-Raros & Cook (1974) described one sample with shorter siphunculi and a more hairy cauda from E. nauseosa as a subspecies, P. packi ssp. brevis, although from their description this seems closer to P. sporadicus.
Pleotrichophorus parilis Corpuz-Raros & Cook Apterae are pale, colour unknown, BL 1.4-2.0 mm. On Artemisia dracunculoides in Minnesota, USA, and recently found on Iva axillaris in Oregon (A. Jensen, pers. com., 2016). Oviparae and alate males occur in September (Corpuz-Raros & Cook 1974).
Pleotrichophorus patonkus (Hottes & Frison) (Fig.5c) Apterae are apple green with darker green spinal and lateral stripes, with hoary appearance due to the numerous capitate dorsal hairs; BL c.1.6-1.9 mm. On Achillea millefolium in central USA (Illinois, Missouri, Tennessee, Kentucky), and apparently widely distributed in North America, as it is also recorded from Achillea spp. in many western states and from Quebec, Canada (A. Jensen, aphidtrek.org and http://www.flickr.com/photos/sandnine/5680760834/in/set-72157626451091245). Aphids collected from Erigeron philadelphicus in Illinois (BMNH collection, leg. V.F. Eastop) may also be this species. Records prior to 1974 including the original description are confused with P. pseudoplatonkus (q.v.); the distinction between these two species still needs further confirmation. Hottes & Frison (1931) described oviparae and alate males collected in October-November, but these may have at least partly been P. pseudopatonkus.
Pleotrichophorus patonkusellus Corpuz-Raros & Cook Colour of apterae in life is unknown, probably pale green with hoary appearance due to the dense capitate hairs; BL 1.1-1.6 mm. On Achillea millefolium and Achillea sp. in Utah, USA (Corpuz-Raros & Cook 1974). Very similar to P. patonkus, apart from the denser and somewhat shorter dorsal hairs, and possibly just a local variant of that species.
Pleotrichophorus persimilis Börner Apterae are pale yellowish red to brownish, appearing wax-powdered; BL 1.9-2.6 mm. On Artemisia spp., especially A. campestris and A. scoparia,in Europe, and east to Iran, Kazakhstan and Pakistan. Oviparae and apterous males occur on A. campestris in September in Germany (Hille Ris Lambers 1953). Green aphids with rather shorter appendages found in Afghanistan on A. turanica are regarded as a subspecies, A. persimilis ssp. afghanensis Narzikulov & Umarov(Daniyarova & Narzikulov 1984), and aphids identified as this subspecies have also now been recorded (but on other Artemisia spp.) from Kazakhstan (Kadyrbekov 2016d, 2017a).
Pleotrichophorus pseudoglandulosus (Palmer) (Fig.14j) Apterae are bluish green, with frosted appearance due to dorsal hairs; BL 1.1-2.0 mm. They live singly on leaves and stems of Artemisia spp. (douglasiana, frigida, ludoviciana, vulgaris) in western USA, and in Canada (Alberta, Manitoba, Saskatchewan). There has been considerable confusion with other species in the literature; records of P. glandulosus from native American Artemisia spp. in western USA are likely to be this species, and most records from Artemisia tridentata are probably referable to the very similar P. decampus. P. intermedius, described from A. frigida in Minnesota (Corpuz-Raros & Cook 1974) is also very similar and possibly a local variant. Oviparae and apterous males (cf. decampus) occur in October-November in Colorado (Palmer 1952) and in September in Manitoba (BMNH collection, leg. A.G. Robinson).
Pleotrichophorus pseudopatonkus Corpuz-Raros & Cook Apterae are whitish green to green, BL 1.3-1.9 mm. On Achillea millefolium and Achillea sp. in north, central and eastern USA, western Canada (New Brunswick) and Mexico (BMNH collection, leg. G. Remaudière). This species is very similar to P. patonkus, but has a shorter, less acute R IV+Vand shorter siphunculi (Corpuz-Raros & Cook 1974). Oviparae and alate males are possibly included in the original description of P. patonkus (Hottes & Frison 1931).
Pleotrichophorus pullus (Gillette & Palmer) (= P. rusticatus (Knowlton & Smith)) (Fig.14d) Apterae are brown with appendages mainly dark brown to black; BL 1.7-2.0 mm. On leaves of Artemisia spp. (cana, longifolia, tridentata) in western USA (Palmer 1952). P. rusticatus (Knowlton & Smith) was regarded as a synonym by Hille Ris Lambers (1969), but Corpuz-Raros & Cook (1974) resurrected it as a good species on the basis of differences in shape of the frontal hairs. After examining types of both, we think this is probably only local variation.
Pleotrichophorus pycnorhysus (Knowlton & Smith) (Fig.21a) Apterae are pale green to deep apple green, somewhat shiny, with long, dark-tipped siphunculi; BL 1.6-2.3 mm. On Chrysothamnus viscidiflorus and Gutierrezia sp. in Utah, USA; also recorded from Idaho, Nevada, Oregon, Washington and Montana, and from Ericameria nauseosa. There is also a record from Artemisia tridentata, but this is unlikely to be a true host plant. A fundatrix was found in May on C. viscidiflorus by A. Jensen (see photo at http://www.flickr.com/photos/sandnine/5692282948/in/set-72157626451091245).
Pleotrichophorus quadritrichus (Knowlton & Smith) (Fig.14c,i) Apterae are bluish green, with greyish cast due to dorsal hairs, and usually with dark siphunculi and cauda (see aphidtrek.org (see influentialpoints.com/Gallery); BL 1.1-2.0 mm. On Artemisia spp. in western USA. The variation in this species was discussed by Corpuz-Raros & Cook (1974), who erected subspecies P. quadritrichus ssp. vulgaris and P. quadtritrichus ssp. pallidus; the latter is a pale form that is keyed separately and resembles P. gnaphalodes. The main distinguishing features for the quadritrichus group are the large, constricted cauda (a feature shared with P. gnaphalodes), and the often strap-like apices of the frontal hairs. Oviparae and alate males occur in October.
Pleotrichophorus remaudierei Leclant Apterae are pale green with a darker spinal stripe; BL 1.2-1.6 mm. Living singly on undersides of leaves of Erigeron acer in the French Alps. Monoecious holocyclic; fundatrices were collected in April (Leclant 1968) .
Pleotrichophorus spatulavillus (Knowlton & Smith) Apterae are pale green, with white frosted appearance; BL 1.4-1.7 mm. On Artemisia tridentata and Artemisia sp. in Idaho and Utah, USA.
Pleotrichophorus sporadicus (Knowlton) Apterae are spindle-shaped, apple green, described as “heavily pruinose above” (presumably an effect of the fan-shaped/spatulate dorsal hairs), with mainly dark siphunculi; BL 2.1-4.1 mm. On leaves and stems of Ericameria nauseosa in western USA. Oviparae were found in October. This is one of a complex of forms described from this host, and distinguished mainly by differences in number and form of dorsal hairs, which are likely to be influenced by environment and by degree of alatiformity. P. neosporadicus Corpuz-Raros and P. packi ssp. brevis are both very close to P. sporadicus, and we could not find reliable distinguishing characters for either of them.
Pleotrichophorus stroudi (Knowlton) Colour of apterae in life was not noted, presumably pale with frosted appearance due to fan-shaped dorsal hairs, and with antennae and siphunculi dark except at their bases (see influentialpoints.com/Gallery); BL 2.0-2.5 mm. Hosts are Ericameria nauseosa and Chrysothamnus viscidiflorus (A. Jensen, aphidtrek.org). In western USA.It belongs to a taxonomically-difficult group of closely-related forms feeding on E. nauseosa (rabbitbrush) in western USA (see packi, sporadicus).
Pleotrichophorus tetradymiae Smith & Knowlton Apterae are “black to blackish green” (?), with black siphunculi; BL 1.5-2.1 mm. On Tetradymia canescens in western USA (Idaho, Oregon).
Pleotrichophorus triangulatus Corpuz-Raros & Cook Apterae are green; BL c.1.7 mm. On Agoseris sp. in western USA (California, Colorado).
Pleotrichophorus tuberculatus Stekolshchikov & Khruleva Apterae are dirty brown-yellow with intensive wax pulverulence both dorsally and ventrally, and dark siphunculi; BL 1.7-2.5 mm. On upper and undersides of apical leaves and on shoot apices of Artemisia tilesii in north-east Siberia (Chukotka). A fundatrix was collected in early July, and oviparae and apterous males were present in early August (Stekolshchikov & Khruleva 2015).
Pleotrichophorus ugamicus Kadyrbekov Apterae are whitish; BL 2.2-2.8 mm. On Artemisia santolinifolia in Kazakhstan (Kadyrbekov 2016a). A member of the glandulosus group.
Pleotrichophorus utensis (Pack & Knowlton) ( = Capitophorus acanthovillus Knowlton & Smith) (Fig.21d) Apterae are green with black siphunculi; BL 1.4-1.9 mm. On Chrysothamnus viscidiflorus and Gutierrezia spp. in western USA (Utah, Nevada,Oregon). P. acanthovillus has been distinguished from P. utensis occurring on the same hosts solely on the basis of differences in shape of dorsal hairs in apterae, and in particular the subapical caudal hair(s), which are said to be pointed in apterae of acanthovillus, and flattened apically in utensis (Corpuz-Raros and Cook, 1974). This character is unreliable as it is related to the degree of alatiformity, alatae having longer and less expanded hairs than apterae.
Pleotrichophorus villosae Robinson Apterae are very pale green, almost silvery (due to dense covering of fan-shaped dorsal hairs); BL 1.6-2.1 mm. On Chrysopsis (= Heterotheca) villosa in Manitoba, Canada. Alatae and other morphs are undescribed.
Pleotrichophorus wasatchii (Knowlton) Apterae are greenish, with apparent pruinosity due to dense covering of fan-shaped hairs; BL 1.6-2.1 mm. On Chrysothamnus and Ericameria spp. in western USA. Records from other host plants and from eastern USA are probably all due to confusion with other species; see Corpuz-Raros & Cook (1974).
Pleotrichophorus xerozoous (Knowlton & Smith) Apterae are green, with antennae dark distally, and siphunculi pale basally and dusky on distal half; BL 1.7-2.5 mm. Alatae are unknown. On Chrysothamnus and Ericameria spp. in Idaho and Utah, USA. This species is closely related to the much commoner P. gregarius, and is possible a seasonal variant, differing in colour in life as well as in characters given in key.
Pleotrichophorus zoomontanus (Knowlton & Smith) Apterae are greyish green, BL 1.7-2.4 mm. On Artemisia tridentata and Artemisia spp. in western USA. It resembles P. longinectarius, especially in having similarly protruding compound eyes, but has a shorter R IV+Vand differences in frontal and dorsal hairs (Corpuz-Raros & Cook, 1974). Alatae are undescribed.
|
Plocamaphis Oestlund |
Aphidinae: Macrosiphini |
A small genus with a rather evenly holarctic distribution, living without host alternation on Salix. They are related to Pterocomma, but less hairy, and have flangeless siphunculi. Most species secrete flocculent wax, and they are not attended by ants. Richards (1966b) revised the genus (which in his concept included Fullawaya) and Wojciechowski (2003) reviewed the palaearctic species. Tang et al. (2015) investigated the use of DNA barcodes for species identification.
Plocamaphis amerinae (Hartig) Apterae are yellowish, greenish or brownish, wax-covered in life, with dark head, antennae and legs; BL 3.0-4.5 mm. On young growth of Salix spp. (alba, purpurea, viminalis), often in large colonies and sometimes causing damage in osier plantations. In north-west and north Europe, west Siberia, and reported also from north-east Siberia (Chukotka; Stekolshchikov & Khruleva 2015) and Kazakhstan (Kadyrbekov 2017a). Possibly also in Canada, if P. martini Richards (q.v.) is this species. Apterous males and oviparae occur in October-November. Populations with very short siphunculi and no dorsal dark markings on a Salix species (probably lapponum) in Sweden were described as a subspecies (P. amerinae borealis) by Ossiannilsson (1959); such forms are also reported from S. viminalis in Poland (Szelegiewicz 1965).
Plocamaphis coreana (Okamoto & Takahashi) (fig. 106C) Apterae are dark yellowish green, covered with white wax; BL c. 3.5 mm. Described attacking branch and stem of an unidentified Salix sp. in Korea (Okamoto & Takahashi 1927; Paik 1965, as Clavigerus salijaponicus). Pashtshenko (1988b) recorded it from S. crassijulis and S. parallelinervis in east Russia (Kamchatca). The life cycle is unknown.
Plocamaphis flocculosa (Weed) (figs 106E,F) Apterae are grey or yellowish grey, densely covered with wax powder; BL 3.1-5.0 mm. Head, legs and antennae are darker, and siphunculi are orange. On trunk and branches of Salix spp. in North America (discolor, lasiolepis) and Europe (borealis, caprea, cinerea, nigricans, repens). Also recorded from Iran (Rezwani et al. 1994) and China. Three subspecies have been distinguished in the very variable European populations (Ossiannilsson 1959), but further work is required to determine whether these are valid taxonomic categories (Heie 1986). The European form known as P. flocculosa ssp. brachysiphon produces oviparae and alate males in October (Stroyan 1955; as P. goernitzi). Kanturski et al. (2016b) described the fundatrix from Poland. P. assetacea Zhang, described from Salix sp. in Tibet (Zhang & Zhong 1981b), may be a synonym (Tang et al. 2015).
Plocamaphis gyirongensis Zhang Apterae and alatae were described from Tibet (Zhang & Zhong 1981b) as a new genus (Radisectaphis), but were transferred to Plocamaphis by Remaudière & Remaudière (1997). The recorded host plant is Malus; presumably this is either an error, or the aphids were vagrants. It is very similar to P. flocculosa, and Wojciechowski (2003) suggested that is should be regarded as a subspecies of that species.
Plocamaphis martini Richards (fig. 106D) Apterae light brown to pink (no wax mentioned); BL 2.8-3.5 mm. On roots of Salix sp(p). in arctic and subarctic Canada (Baffin Island, Quebec). Apterous males and oviparae occur in mid-August (Richards 1963). Possibly conspecific with P. amerinae.
|
Polytrichaphis Miyazaki |
Aphidinae: Macrosiphini |
Two east Asian species on Illiciaceae, with numerous dorsal hairs and thick, tapering siphunculi resembling Indiaphis,but first tarsal segments have two hairs displaced from apex, the tergum is membranous and the eyes have distinct ocular tubercles. Alatae have dark-bordered wing veins and secondary rhinaria on ANT III-IV or III-V.
Polytrichaphis fragilis Miyazaki Apterae are yellow to pale-yellowish green with dark brown siphunculi and a pale cauda; BL c.1.6-1.8 mm. On undersides of leaves of Illicium anisatum in Japan(Miyazaki 1971).
Polytrichaphis illicius Qiao & Zhang Apterae are black; BL c.1.6-1.8 mm. On Illicium verum in China (Qiao & Zhang 2001d).
|
Prociphilus Koch |
Eriosomatinae: Pemphigini |
About 45 species ranging in body size from rather small to very large, usually lacking siphunculi and with well-developed wax glands, even in fundatrices. Their host plant associations are unusual, in that they have apparently transferred their sexual generation in relatively recent times from Populus to various more advanced “primary” hosts, e.g. in Rosaceae, Caprifoliaceae and Oleaceae, while mostly retaining conifer roots as secondary hosts for the parthenogenetic generations. The species are fairly evenly distributed through the northern hemisphere; about 20 in North America, 7 in Europe, 3 in central Asia and 15 in east Asia. The life cycles of many species are unknown. Accounts are available for north-west Europe (Heie 1980b), the UK (Blackman et al. 2019b), Spain and Portugal (Nieto Nafría et al. 2002a), Central Asia (Narzikulov 1964), India (A.K. Ghosh 1984), Japan (Inouye 1956), China (Tao 1970, G. Zhang et al. 1993c, G. Zhang et al. 1999c) and North America (Smith 1974a). Our concept of Prociphilus includes Stagona, Paraprociphilus (with migration from Acer to Alnus) and we also include as subgenera Pulvius, Neoparacletus and Meliarhizophagus, each with a single species, which all appear to be relatively recent biological deviations from the main evolutionary lineage of Prociphilus.
Prociphilus americanus (Walker) Host-alternating between Fraxinus spp. and roots of Abies (balsamea, grandis, procera) in North America. The type specimen is an alate sexupara from Nova Scotia (Doncaster 1961); prior to Doncaster’s redescription this aphid was called P. fraxinidipetalae and/or P. venafuscus. Early records from roots of Pseudotsuga are probably in error (Smith 1969), and records from other trees (eg. Aesculus) are probably of vagrant individuals. Fundatrices on ash are dark brown to black, globose, forming early spring colonies on in curled terminal leaves of new growth (Essig 1911) or on twigs (Patch 1909); populations may also develop on suckers and at base of trunk “at quite a depth underground” (Essig). Emigrant alatae maturing from late April to early June are very dark green to black with dusky, bluish-tinged wings and long bluish white wax flocks extending posteriorly; BL 2.8-4.8 mm. They found wax-covered colonies on Abies roots; a redescription of the apterous exule (BL 1.4-2.3 mm) was provided by Pike et al. (2012). Sexuparae return to produce dwarf sexual morphs on trunk of Fraxinus in late August-October (Smith 1969). Anholocyclic overwintering may also occur on Abies roots (Zak 1965). Patch (1909) described the sexual morphs (as venafuscus) and recorded that they were also deposited in large numbers on Syringa, which may be an alternative primary host. This is probably the aphid referred to as “Prociphilus californicus” in some western North American publications (see Halbert & Blackman 2004).
Prociphilus aurus Zhang & Qiao Only the fundatrix is described, and its appearance in life is unknown; BL c. 2.9 mm. On Malus sieversii in Xinjiang, China (G. Zhang & Qiao 1997c).
Prociphilus (Paraprociphilus) baicalensis (Cholodkovsky) Apterae brownish, secreting copious white wax wool: BL c. 3.1 mm. On undersides of leaves and shoots of Alnus spp., across Asia from the Urals to Japan. Apparently entirely anholocyclic on Alnus, although sexuparae are produced in autumn (Mordvilko 1935). 2n=12*.
Prociphilus bumeliae (Schrank) Host-alternating between Oleaceae (usually Fraxinus excelsior, sometimes Ligustrum vulgare, Syringa vulgaris) and roots of Abies. Fundatrices are brown, covered with white wax-wool, forming loose “leaf-nests” on primary host in spring. Spring colonies also form on bark of stems and branches (see bladmineerders.nl). All their progeny are alate, with blackish brown head and thorax spotted with white wax, and light brown to yellowish red abdomen (Heie 1980b); BL 3.8-5.5 mm. They migrate in May-June to found wax-covered colonies on Abies roots. (There is a possibility that they also live attended by ants on roots of Picea abies –see under P. fraxini). Sexuparae return to Fraxinus in late August-September (Mordvilko 1935). Widely distributed in north-west, north, central and eastern Europe. Also recorded from east Asia, but its presence there needs confirmation as most of these records should probably be referred to P. oriens. [Alatae of Prociphilus “sp. B” on Fraxinus floribunda in northern India (A.K. Ghosh et al. 1970) key to bumeliae, except that they lack discernible dorsal cephalic wax glands and have many more secondary rhinaria (45-48, compared with 16-24) on ANT III.]
Prociphilus carolinensis Smith Apterae are oval, presumably wax-covered in life, with protruding dark cauda and anal plate; BL c. 2.5 mm. They live on the roots of Pinus spp. (including P. taeda), 3-4 feet below ground, attended by ants. Sexuparae (BL 2.8-3.3 mm) emerge in October-December, migrating to an unknown primary host. Sexuales are deposited and eggs laid on the bark of various trees, especially Malus, but apparently do not hatch (Smith 1969). In USA (North Carolina) and Canada (Manitoba).
Prociphilus caryae (Fitch) (=alnifoliae Williams) Spring populations (fundatrices + immature alatae) cup or curl the leaves of Amelanchier spp. and turn them yellow or whitish. (Records from Crataegus are probably due to misidentification, particularly confusion with P. corrugatans.) Emigrant alatae (BL 2.7-3.3 mm) fly in June to colonise roots of Pinus sp(p), sexuparae returning to Amelanchier in September-October. In western USA and Canada (British Columbia). (No field observations have been made of these western populations on pine roots, but Smith (1969) obtained colonies readily on Pinus strobus in the laboratory, and sexuparae collected on P. monticola in British Columbia (BMNH collection, leg G. Shrimpton) appear to be this species.) Similar aphids in eastern USA are regarded as a subspecies:-
Prociphilus caryae ssp. fitchii Baker & Davidson Spring populations (fundatrix + immature alatae) cause cupping and curling of leaves of Malus spp. (domestica, angustifolia) and Amelanchier spp. (see influentialpoints.com/Gallery). Emigrant alatae (BL 1.8-3.3 mm) migrate in June to found ant-attended colonies on roots of Pinus strobus. Sexuparae return to the primary hosts in October-November where they deposit sexuales in bark crevices near base of tree or in nearby leaf litter (Smith 1969). Widely distributed in eastern North America. Parthenogenetic populations also persist throughout the year on white pine roots. A closely-related form, P. caryae ssp. arbutifoliae, forms spring populations on Photinia (= Heteromeles) arbutifolia in California, from which alatae (BL 2.9-4.1 mm) migrate to an unknown secondary host, probably pine roots. [Records of P. caryae fitchii on Lonicera in India (A.K. Ghosh et al. 1981, A.K. Ghosh 1984b) apply to P. himalayensis.]
Prociphilus chaenomelis Lee, Seo & Hwang Apterae (fundatrices) are undescribed, alatae are dark brown producing copious wax; BL 2.1-2.8 mm. In rolled leaves of Chaenomeles sinensis in Korea. Presumably heteroecious holocyclic, migrating to an unknown secondary host.
Prociphilus clerodendri Okamoto & Takahashi Apterae are not described. Alatae have head, eyes, antennae and mesothorax black, legs black and reddish brown, and abdomen yellowish green with a reddish tinge, secreting white cottony wax; BL c. 4.5 mm. On Clerodendron trichotomum in Korea.
Prociphilus cornifoliae Singh, Das & Raychaudhuri Apterae (BL 4.2 mm, colour in life unrecorded) were collected on roots of Cornus sp. in April in Manipur, India (Singh et al. 1977; habitat information from A.K. Ghosh 1984b). Alate specimens and immatures were also collected, both from the same host, locality and date, and from an unidentified plant in Nepal. Life cycle is unknown.
Prociphilus (Neoparacletus) corrugatans (Sirrine) Spring populations cause curling and cupping of leaves of Crataegus spp.; infested leaves turn pale whitish, often with a reddish tinge at the edge. Fundatrices are yellowish green with flocculent wax, with greenish immatures that develop into emigrant alatae (BL 1.9-2.9 mm, with brownish abdomen). Migration occurs in May-June to roots of Cyperaceae, where the apterous exules are milky white, sometimes tinged with pale brown, entirely covered with wax powder; BL 2.9-3.8 mm. Large colonies are formed on Juncus spp. and populations may persist throughout the year. It is also recorded from Carex (Strom 1942, as Neoparacletus caricis). Sexuparae returning to Crataegus in autumn often deposit sexuales on Malus, but apparently these do not produce spring populations (Smith 1972a). Records from Amelanchier are possibly all due to confusion with P. caryae. Widely distributed in North America. [Dark brown or reddish curled leaves on Crataegus tomentosa, noted in the original description of P. corrugatans (Sirrine 1894), are considered by Smith (1972a) to be due to feeding by another Prociphilus sp., possibly the undescribed primary host generations of P. erigeronensis.]
Prociphilus crataegicola Shinji Described from Crataegus sp. in June in Japan; alata has light greenish abdomen, and antennae with secondary rhinaria distributed III 22, IV 7, V 5 (Shinji 1922a). Other morphs and life cycle unknown, except that alatae collected from pine roots in Taiwan and Japan by Tao (1970), as “Prociphilus pini n. sp.” could be the sexuparae of this species. Possibly crataegicola is a synonym of pini Burmeister; alatae (BL 3.2 mm) collected on “apple” in Japan and identified by Takahashi as crataegicola (BMNH collection, leg. T. Tanaka) have secondary rhinaria distributed III 24-29, IV 6-8, V 6-8, and closely resemble European P. pini. [P. sasakii Monzen, described from Malus domestica was synonymised with crataegicola by Shinji (1941), but this seems doubtful as Monzen’s species had siphunculi (unusual in Prociphilus), and less rhinaria on III.]
Prociphilus (Stagona) crataegistrobi Smith Fundatrices and their progeny in spring curl leaves of Crataegus sp., which remain green but may tend to fade. Their progeny are all alate, BL 2.2-2.5 mm, migrating in June to found colonies on roots of Pinus strobus. The apterous exules live in indvidual wax-lined cells associated with small roots and mycorrhizae (Smith 1969). Sexuparae return to trunks of Crataegus in October-December, but pine root-feeding populations also persist through winter. In USA; only known from North Carolina.
Prociphilus (Stagona) dilonicerae Zhang Apterae (fundatrices) are undescribed; BL of alata (fundatrigenia) is c.3.6 mm. On Lonicera sp. in Tibet. Secondary host is unknown.
Prociphilus emeiensis Zhang Described from two alatae (BL c. 4.0 mm) collected in April on Fraxinus chinensis in Sichuan, China (G. Zhang & Qiao 1997c). Appearance in life and biology are unknown.
Prociphilus erigeronensis (Thomas) Plate 2h Apterae (exules) are white to pale yellow with darker head, antennae, legs, and end of abdomen, secreting white wax; BL 1.7-2.3 mm. In ant-attended colonies on roots of many plants, especially Compositae (Ambrosia, Aster, Cichorium, Erigeron, Lactuca, Solidago, Tragopogon), Poaceae (Agrostis, Muhlenbergia, Poa, Triticum) and Leguminosae (Arachis, Phaseolus, Trifolium). Widely distributed in USA, and also recorded from Ontario, Canada, and from Puerto Rico. Predominantly anholocyclic; Smith (1974) redescribed this species, including alate sexuparae, and discussed a possible partial holocycle with Crataegus (or Amelanchier) as primary host. The name is possibly being applied to more than one species.
Prociphilus formosanus Takahashi Dense spring colonies of this species distort the leaves of Fraxinus sp. (“insularis“) in Taiwan (Takahashi 1935). Alatae (collected in May) have a dark green abdomen, and BL c. 4 mm; antennae with secondary rhinaria distributed III 32-39, IV 7-11, V 7-9, VI 0. Immatures yellow with cottony wax on abdomen. Other morphs are unknown. Alatae were also collected on Picea morrisonicola, which is therefore thought to be the secondary host, but confirmation of this seems necessary.
Prociphilus fraxini (Fabricius) Host-alternating between Fraxinus excelsior and roots of Abies, and probably also Picea (see below). Fundatrices feed at bases of new shoots of ash, giving rise to large colonies of immature alatae which develop to maturity in dense leaf-nests, often high up in trees (Heie 1980b). Alatae (BL 3.3-5.1 mm) fly in late June-July to found wax-covered colonies on Abies roots, the apterous exules being almost indistinguishable from those of P. bumeliae. Sexuparae return to Fraxinus in October-November. The male genitalia were described and illustrated by Wieczorek et al. (2012). [Immature apterous exules identified as P. fraxini have now been observed attended by ants (Lasius brunneus and Prenolepis nitens) at bases of trunks of Picea abies, andthesemay have overwintered parthenogenetically deep underground on spruce roots in symbiotic association with specialised ants such as P. nitens (Purkart et al. 2019).] P. fraxini is widespread in Europe, and also recorded from Iran; specimens from Turkey on Fraxinus oxycarpa (BMNH collection, leg. H. Canakçioglu) are possibly an undescribed, closely-related species. Also recorded from China (G. Zhang et al. 1993c).
Prociphilus (Meliarhizophagus) fraxinifolii (Riley) Living all-year-round on American Fraxinus spp. Yellow-green to pale green fundatrices and their progeny (both apterae and alatae) live in curled and clumped leaves at tips of twigs throughout the growing season (see influentialpoints.com/Gallery). Alatae (BL 1.4-2.8 mm, with yellow-green abdomen) are produced in April-October. Ant-attended colonies also occur all-year-round on Fraxinus roots. Alate sexuparae occur in October-November (Smith 1974a). Common and widely distributed in USA; also in Canada (British Columbia, Manitoba) and Mexico, and introduced to Chile (Carrillo 1977), South Africa (on planted F. pennsylvanica and F. velutina; Müller & Schöll 1958, and BMNH collection); also to Europe, where a colony was found on a planted F. pennsylvanica in Hungary (Remaudière & Ripka 2003), and it has subsequently spread to Serbia (Petrović-Obradović et al. 2007), Bulgaria (Trenchev & Trencheva 2009), Spain (Pérez Hidalgo & Mier Durante 2012), UK (Baker & Martin 2011), Poland (Halaj et al. 2016), Iran (Tajmiri et al. 2016), Kazakhstan (Kadyrbekov 2017a) and Romania (Oleniciet al. 2018). In 2015 it was found for the first time on the native European ash F. excelsior at four localities in Poland (Halaj & Osiadacz 2017). It has also recently been reported from Beijing, China (Yu et al. 2015). Carver (1980) described an aphelinid parasitoid on P. fraxinifolii in Iowa, and Brudrett & Kendrick (1987) studied symbiosis between root-feeding populations and a bolete fungus. 2n=20 (Robinson & Chen 1969) or 2n=22 (Blackman & Eastop 1994).
Prociphilus gambosae Zhang & Zhang Apterae are greyish green, secreting long wax filaments; BL c. 2.4 mm. On Syzygium caryophyllus (= aromaticum) in Hebei, China (G. Zhang et al. 1993c). Other morphs, biology and life cycle are unknown.
Prociphilus (?Neoparacletus) ghanii Hille Ris Lambers Apterae are broadly oval, appearance in life unknown; BL 1.3-1.7 mm. Collected on Salix wallichiana in Pakistan (Hille Ris Lambers 1973b). Other morphs, biology and life cycle are unknown.
Prociphilus himalayaensis Chakrabarti Described from sexuparae (BL 2.6-3.4 mm, wax-covered) collected on roots of Pinus excelsa (=griffithii) in Himachal Pradesh, India (Chakrabarti 1976a). This aphid was originally placed in subgenus Stagona, although the pterostigma is not elongated, perhaps because the apterous exules described as paratypes (from an unidentified host) have a tibial chaetotaxy like that of P. (S.) crataegistrobi, but there is no real evidence that these are the same species as the sexuparae from pine roots. Banerjee & Chakrabarti (1993) confirmed experimentally the suspected (Chakrabarti et al. 1985) migration from rolled-leaf galls on Lonicera quinquelocularis. The spring forms on Lonicera had earlier been recorded as P. caryae fitchii by A.K. Ghosh (1984b). The apterae (fundatrices) are pale brown; BL 2.2-3.3 mm. All progeny are alate (BL 2.3-3.0 mm), migrating in May-June to roots of P. excelsa. Alate sexuparae (BL 2.6-3.4 mm) return to Lonicera in November-December (Banerjee & Chakrabarti 1993). Samples have been seen from the Punjab, Himachal Pradesh, Kashmir and Pakistan. Rather similar spring migrants but with more rhinaria were collected from Eriobotrya petiolata in West Bengal (BMNH collection, leg. A.N. Basu), so there remains the possibility that two species with different primary hosts are being confused. There are apterae of other undescribed species on roots of Pinus griffithii in the Himalayas, and the group merits further investigation. P. trinusZhang, collected from L. japonica in China, is probably a synonym on the basis of the published description. The distinction between P. himalayensis and the Central Asian species P. umarovi is also unclear.
Prociphilus (Stagona) konoi Hori Host-alternating between Lonicera spp. and Picea. Fundatrices are yellowish green, BL 2.5-3.0 mm, curling terminal leaves of Lonicera longitudinally. All their progeny are alate (BL 2.5-2.8 mm, with greenish brown abdomen) and migrate in June-July to found colonies on roots of Picea glehnii (Hori 1938a; requires experimental confirmation). Sexuparae (with secondary rhinaria distributed ANT III 15-16, IV 8-9, V 7-8, VI 3-4) return to Lonicera in October-November. In Japan and east Siberia (Pashtshenko 1988a). 2n=18.
Prociphilus kuwanai Monzen(= orientalis Mordvilko 1929) Feeding by the fundatrix in spring on Pyrus spp. (and occasionally Malus) causes leaf to curl downwards forming a bag-like gall, coloured yellowish green tinged with red. Alatae (BL 2.6-2.9 mm, with a yellowish green abdomen) fly in late June-early July. The secondary host is uncertain; roots of Larix according to Shinji (1932; possibly due to confusion with another species), or more probably of Rumex spp. (Mordvilko 1935, Higuchi & Miyazaki 1969). Hori (1938a) also noted that the secondary host “may probably be Rumex“. Sexuparae occur in October-November, but have not been described. In Japan, Korea and east Siberia.
Prociphilus (?Stagona) laricis Shinji Only alate sexuparae are described, BL c. 1.6 mm, found swarming out of the ground along the roots and trunk of Larix kaempferi in Japan in late October-early November (Shinii 1941). The primary host is unknown.
Prociphilus ligustrifoliae (Tseng & Tao) Spring populations occur in curled leaves of Ligustrum japonicum in China (Tseng & Tao 1938), and are also recorded from L. lucidum in Japan (Wieczorek et al. 2012). The globular yellowish white fundatrices, covered in white cottony wax, develop on the buds in March-April and cause economic damage to trees used in rearing the coccid Ericerus pela for commercial wax production in Sichuan (Zhang 1981). The alatae produced in the leaf-nests, have BL c. 3.75 mm, dark-bordered wing veins, a pale green abdomen, and secondary rhinaria distributed III 32-36, IV 13-16, V 12-16, VI (0-)1-7. They presumably fly to an unknown secondary host. This species is closely related to, and possibly synonymous with, P. osmanthae.
Prociphilus longianus Smith The appearance of the apterae (exules) in life is unrecorded, presumably secreting white wax, with conspicuous dark sclerotic tubular extension of anal plate around cauda; BL 2.7-3.3 mm. The host is uncertain; it was recorded from a “gall on Conophilus americanus”(presumably Conopholis americana), also from base of stem of a “garden geranium”, roots of an unidentified grass, and from ants’ nests.
Prociphilus (Stagona?) lonicerae Shinji Apterae (fundatrices) are green; BL not recorded. Rolling young leaves of Lonicera japonica in Japan. All progeny are alate (BL c.2.1 mm), migrating in early June to an unknown secondary host.
Prociphilus (Paraprociphilus) mexicanus Remaudière & Munoz Viveros Apterae are dark-coloured, entirely covered with a dense waxy secretion, with long wax filaments projecting posteriorly; BL 2.9-3.6 mm. On undersides of leaves of Alnus firmifolia in Mexico (Remaudière & Munoz Viveros 1986). Sexuparae are produced in autumn, and there is at least a partial migration to Acer negundo, from which fundatrices and emigrant alatae were described by Remaudière & Munoz Viveros (1992b).
Prociphilus micheliae Hille Ris Lambers Only spring migrant alatae are described; appearance in life unknown, BL 3.3-4.0 mm, secondary rhinaria distributed III 28-35, IV 8-13, V 0, VI 0. In curled leaves of Michelia (= Magnolia) champaca in northern India (Kashmir, West Bengal, Uttar Pradesh), becoming mature in April-June (BMNH collection). Other morphs amd life cycle are unknown. 2n=14 (Kar et al. 1990).
Prociphilus oleae Leach ex Risso Fundatrices (BL c.3.2 mm) and their progeny form compact spring colonies covered in white wax wool on shoots near base of trunk of Olea europea. It is also recorded from Phillyrea latifolia Unusually for a Prociphilus, no leaf deformation is caused. Progeny of the fundatrix are all alate, BL 3.7-4.0 mm with smoky yellowish abdomen, migrating in May. The life cycle is unknown, but apterae described from the roots of Vitis vinifera in Sicily (Barbagallo & Stroyan 1982) could be the secondary host form of this species. In Mediterranean area (France, Italy, Greece, Turkey). Roberti & Monaco (1987) gave an account of this aphid in Italy.
Prociphilus oriens Mordvilko Fundatrices and their progeny feed on young growth of primary hosts in spring causing curling and clustering of leaves into leaf-nest galls (see http://www.agr.hokudai.ac.jp/systent/akimoto/gall/P.%20oriens.jpg). Typical primary hosts are Fraxinus spp., but there are records of spring populations on a remarkable range of other plants, not only in Oleaceae (Ligustrum, Lonicera, Syringa) but including Rosaceae (Malus, Prunus, Sorbus) as well as Salix, Tilia and Ulmus (Kono 1940). The progeny of the fundatrix are all alatae, BL 3.5-5.5 mm, with yellowish brown to dull reddish green abdomen covered with white wax; migrating in June-early July to roots of Abies sachalinensis (incl. var. mayriana). Records from Picea roots need confirmation. Return migration of sexuparae occurs in September-October (Hori 1938a). Recorded from India, China, east Siberia, Korea and Japan (type locality). Yamaguchi (1985) and Li & Akimoto (2017) studied sex ratio variation in this species, and Akimoto (2006) investigated the effects of in-breeding.
{kind=link}
Prociphilus osmanthae Essig & Kuwana Apterae in curled-leaf pseudogalls on Osmanthus spp. in spring are greyish green, wax-covered (see influentialpoints.com/Gallery); BL c. 1.8 mm. Emigrant alatae have BL 3.3-4.0 mm, wax-covered olive-green abdomen, and dark-bordered forewing veins (Essig & Kuwana 1918, Moritsu 1983). Secondary rhinaria of paratypes are distributed III 27-31, IV 9-12, V 8-12, VI 0. They migrate in late May to an unknown secondary host. In Japan, and reported to occur in Meghalaya, India (A.K. Ghosh & Raychaudhuri 1968c, on an unidentified host, and Khuda-Bukhsh & Kar 1990, on Croton sp.). It has recently been introduced to Hungary (Ripka et al. 2020). A.K. Ghosh (1984b) included various Japanese specimens collected by Takahashi in his redescription of osmanthae; emigrant alatae from Ligustrum, apterous exules from Michelia, and alate sexuparae (with an anocaudal process) from Osmanthus. It is not certain that all these forms are the same species. P. ligustrifoliae also has dark-bordered wing veins and may be a synonym, although alatae from galls have more rhinaria, including some on ANT VI. 2n=18 (for aphids collected on Croton; Khuda-Bukhsh & Kar 1990).
Prociphilus pergandei Smith Fundatrices curl leaflets of Fraxinus americana in eastern USA. Progeny are all alate, BL 2.7-3.2 mm, covered with long flocculent wax secretion, flying in June to an unknown secondary host (Smith 1974a).
Prociphilus (Stagona) piceaerubensis Smith Apterae are white to pale yellowish with a tuft of wax on abdomen; BL 0.9-1.2 mm. On roots of Picea rubens, inhabiting individual wax-lined cells associated with mycorrhizae, especially in humus or where roots have penetrated decaying logs. Sexuparae occur in September-October. Described from North Carolina, USA (Smith 1969); sexuparae which may be this species have also been trapped in Germany and England (BMNH collection), and it is perhaps the form described as xylostei “forma minima” by Tüllgren (1909).
Prociphilus (Stagona) pini (Burmeister) Plate 6a, b Fundatrices and their progeny curl and yellow the leaves of Crataegus spp. in spring. Progeny are all alate, BL 1.7-3.1 mm, with light green to greyish green abdomen, flying in June-August to found colonies of small, wax-covered apterae on fine roots of Pinus sylvestris and its close relatives; also recorded on P. contorta in UK (Carter & Danielsson 1993) and from P. sibirica in Kazakhstan (Kadyrbekov 2017a). Sexuparae (BL 1.6-2.3 mm; (see influentialpoints.com/Gallery) return to Crataegus in October-November, but populations also persist throughout the year on Pinus roots. Tambs-Lyche & Heie (1994) recorded (vagrant?) P. pini from Malus domestica. In north-west and central Europe, and Central Asia (Kazakhstan; Kadyrbekov 2017a). Smith (1969) gave distinguishing characters from the closely-related North American species crataegistrobi. 2n=16.
Prociphilus piniradicivorus Smith Apparently living all-year-round on Pinus spp. (Smith 1969). Apterae (presumably wax-covered in life, BL c. 2 mm, with protruding dark cauda and anal plate) live on roots of Pinus spp., attended by ants and often in association with the very similar but host-alternating species P. carolinensis. Sexuparae (BL 2.9-3.3 mm) are produced in October-November and fly only a short distance, depositing sexual morphs on trunks of nearby pine trees. Colonies were experimentally reared from fundatrices that hatched from eggs transferred to seedling P. taeda (Smith 1969). In USA (North Carolina, Maryland, Indiana).
Prociphilus (Pulvius) probosceus (Sanborn) Living all-year-round on Fraxinus. Fundatrices and their developing alate progeny curl and distort leaves and terminal growth of Fraxinus spp. in spring. More commonly found are the very large apterae (BL 4.5-5.3 mm), in ant shelters at bases of water shoots and trunks of young trees, at or below soil level (Smith & Stroyan 1972). Sexuparae are produced in October-November, but underground colonies persist throughout the year. In eastern North America.
Prociphilus sasakii Monzen Described from Malus domestica in Japan (Monzen 1927, 1929). Large, globular fundatrices (BL c. 3.3 mm) and their developing alate progeny, covered with white woolly wax, cause apple leaves to become curled, bloated and yellowed in spring. Alatae, BL c. 3.1 mm with pale green abdomen, and secondary rhinaria distributed ANT III 15-18, IV 5-8, V 5-7, VI 0, migrate to an unknown secondary host. (See also discussion of this species under P. crataegicola.)
Prociphilus taxus Ghosh, Chakrabarti, Chowdhuri & Raychaudhuri Only known from the alate sexupara, collected on roots of Taxus baccata in Himachal Pradesh, India (A.K. Ghosh et al. 1969); BL 2.9-3.5 mm, secondary rhinaria distributed ANT III 18-22, IV 8-10, V 6-10, VI2-5). Sexuparae have also been trapped in Pakistan (Naumann-Etienne & Remaudière 1995). The anal plate projects as a collar around the cauda; a similar anocaudal process is known in sexuparae of certain other Prociphilus (keyed by A.K. Ghosh 1984, p.287). The life cycle is unknown.
Prociphilus (Paraprociphilus) tessellatus (Fitch) (fig.18C) Brownish aphids living beneath a copious secretion of white woolly wax. Fundatrices and their alate progeny develop on undersides of leaves of Acer saccharinum in May-July, causing slight leaf-curl. Migration occurs to Alnus, where colonies of large brown wax-covered apterae (BL 3.6-4.4 mm) are formed along the branches (see influentialpoints.com/Gallery). Alate sexuparae return to Acer in September, but anholocyclic overwintering also occurs under fallen leaves or in loose earth at base of alder trees (Patch 1911, Mordvilko 1935). Widely distributed in North America. A syrphid (Chrysopa slossonae) lays its eggs specifically in relation to colonies of P. tessellatus (Bristow 1988). 2n=6 (Blackman & Eastop 1994).
Prociphilus umarovi Narzikulov Apterae (fundatrices) are green; BL 2.0-2.3 mm. Curling leaves of Lonicera spp. in spring. All progeny are alate (BL 2.2-3.6 mm), migrating in June to conifer roots (Abies sibirica, Picea schrenkiana; Kadyrbekov 2017a). In Central Asia (Kazakhstan, Tajikistan). It is possible that P. himalayensis (q.v.) is a synonym. Gabrid (1989) described a much larger aphid from Lonicera spp. in Kyrgyzstan (BL of fundatrices 3.8-4.0 mm, BL of alatae 3.8-4.3 mm), with 14-20 rhinaria on ANT III, as Prociphilus sp. aff. alatavicus Juchnevitch. P. alatavicus is currently regarded as a synonym of P. umarovi.]
Prociphilus ushikoroshi Shinji Fundatrices and their developing all-alate progeny feed on leaves of Photinia (=Pourthiaca) villosa in spring, causing edges of leaves to curl towards underside and become thickened and yellowish green (Monzen 1929; as P. pourthiacae). Alatae (BL c. 2.1 mm, with yellowish green abdomen covered in white woolly wax, and secondary rhinaria distributed ANT III 10-14, IV 4-7, V 1-4, VI 0-1) migrate in June. The secondary host is uncertain, possibly roots of Larix kaempferi (Shinji 1932, Moritsu 1983). Sexuparae return to deposit sexual morphs on trunk of Photinia in October (Monzen). In Japan and Korea. [In India, alatae with many more secondary rhinaria (III 35-36, IV 9-10, V 10-11, VI 0-2) have been collected on Photinia notaniana (A.K. Ghosh et al. 1970; Prociphilus “sp. A”).]
Prociphilus (Stagona) xylostei (De Geer) (Fig.38a) Fundatrices (BL 3.0-4.6 mm) and their developing alate progeny live in copious wax on shrubby species of Lonicera in spring, causing leaves to curl and become spotted with yellow (Mordvilko 1935, Heie 1980b, and see bladmineerders.nl). Alatae (BL 2.3-3.7 mm, with pale green abdomen and white woolly wax (see influentialpoints.com/Gallery)) complete their development on the twigs in late June-July and migrate to found colonies of small, wax-covered apterae (BL 0.8-1.2 mm) on fine roots of conifers. Picea abies is the recorded secondary host in Europe, and P. obovata in Central Asia. In North America it is reported from Abies grandis, but there are some discrepancies between the redescription of apterae from Abies roots by Pike et al. (2012) and the descriptions of European material from roots of Picea. Sexuparae return to Lonicera in October-November. Widely distributed in Europe, and in Central Asia (Kazakhstan; Kadyrbekov 2017a), India (Uttar Pradesh, Kashmir), China (G. Zhang et al. 1993c; recorded from Abelia biflora), and introduced to North America. Differences from P. piceaerubensis were discussed by Smith (1969). Populations in east Siberia are regarded as a subspecies, P. xylostei ussuricus Pashtshenko 1988a (= P. xylostei ssp. orientalis Mordvilko 1935; nec orientalis Mordvilko 1929). 2n=10 (samples from Europe and India).
|
Protaphis Börner |
Aphidinae: Aphidini |
About 50 nominal species closely related to Aphis, but with short appendages and other morphological features adapted to feeding at stem bases and on roots of plants in close association with ants. Most species feed on Compositae/Asteraceae. It is a taxonomically difficult group, many very similar species having been described, and there is a danger that morphological resemblance could be due to convergence associated with feeding position and ant attendance. Most nominal species are in Europe and Central Asia, but three were described from Africa, and six are North American. The North American and Eurasian/African species show some differences, but there is now molecular evidence that they may be sister groups clustering separately from Aphis (Lagos et al. 2014), and therefore justifying the treatment of Protaphis as a distinct genus. However this still needs to be confirmed by more comprehensive DNA studies of Palaearctic species. Kadyrbekov (2001b) reviewed and keyed the central Asian species, and Nieto Nafría et al. (2005a) reviewed those on the Iberian peninsula.
Protaphis afghanica (Narzikulov & Umarov) Apterae are green with darker dorsal markings; BL c. 1.6 mm. On leaves and shoot apices of Artemisia turanica in Afghanistan.
Protaphis alhagii Juchnevitch Apterae are dark green, wax dusted; BL c.1.7 mm. In ant-attended colonies at stem bases and on roots of Alhagi spp.in southern Kazakhstan.
Protaphis ancathiae Kadyrbekov Apterae are greenish marked with dark brown, siphunculi black; BL 1.46-1.7 mm. On roots of Ancathia igniaria in Kazakhstan (Kadyrbekov 2001b); subsequently recorded from Cirsium incanum (= arvense) (Kadyrbekov & Aoitzhanova 2005).
Protaphis anthemiae Ivanoskaya Apterae are dark greenish, with slight grey wax film, siphunculi black and cauda green; BL 1.7-1.8 mm. On roots of an Anthemis sp. attended by ants. Described from Azerbaijan, and subsequently recorded from Kazakhstan (Kadyrbekov 2017a).
Protaphis anthemidis (Börner) Colour of apterae in life unrecorded; BL 1.4-2.0 mm. On roots of Anthemis tinctoria in central and eastern Europe. It is also recorded from Iran (Rezwani et al. 1994) and Kazakhstan (on Achillea millefolium; Kadyrbekov 2011b). Monoecious holocyclic with apterous males (Tashev 1964a). Very similar to P. carlinae and P. dudichi; experimental work is needed to determine whether or not these are all distinct species.
Protaphis anuraphoides (Nevsky) Plate 7d Apterae are green to dark green, coated with mealy wax; BL 1.5-2.1 mm. Alatae have secondary rhinaria distributed III 5-10, IV 0-2. On upper sides of leaves, stems and flowerheads, often in large numbers. On plants in several composite genera of tribe Cardueae (e.g. Carthamus, Carduus, Cirsium, Cousinia, Onopordum). Records from genera in other subfamilies of Compositae/Asteraceae are likely to be misidentifications. In Ukraine, southern Russia and south-west and Central Asia. Kadyrbekov (2001b) redescribed this species; it is very similar to P. carthami.
Protaphis aralensis Kadyrbekov Apterae are greenish, with dark brown head, dorsal markings, siphunculi and cauda, covered with a slight grey wax film; BL 1.7-2.0 mm. On Scorzonera parviflora, at base of stem, attended by ants.In southern Kazakhstan (Kadyrbekov 2001b). Subsequently recorded from Tragopogon dubius (Kadyrbekov & Aoitzhanova 2005).
Protaphis astrakhanica Stekolshchikov Apterae are greyish green, lightly wax-powdered; BL 1.9-2.4 mm. Alatae have secondary rhinaria distributed III 4-8, IV 0-1, V 0. At stem bases near root collars of Lactuca tatarica (= Mulgedium tataricum) in southern Russia (Astrakhan province), covered by soil and attended by ants (Stekolshchikov 2005). The life cycle is unknown, but probably monoecious holocyclic.
Protaphis betpakdalensis Kadyrbekov Apterae are dark greenish variably marked dorsally with dark brown, and a slight grey film of wax; BL 1.9-2.2 mm. On roots of Taraxacum monochlamydeum, attended by ants (Kadyrbekov 2001b). In Kazakhstan.
Protaphis bimacula (Narzikulov & Umarov) Apterae are dark green, wax-dusted, with dark brown head, antennae, apices of femora and tibiae, and a pair of black patches on the dorsal abdomen between the short, dark siphunculi; BL c. 1.45 mm. Colonising apices of stems of Campanula sp. in Afghanistan.
Protaphis carlinae (Börner) Apterae are dark blue-green; BL 1.5-2.1 mm. On rosette leaves of Carlina spp. in central and eastern Europe, and now recorded from southern England (see influentialpoints.com/Gallery). Monoecious holocyclic. Tashev (1964a) redescribed this species, including oviparae; it is very similar to, and possibly synonymous with, P. dudichi.
Protaphis carthami (Das) Apterae are dark green or reddish brown coated with greyish white wax, and often there are a pair of dark spots on dorsal abdomen just anterior to the siphunculi; BL 1.6-2.3 mm. Apterae have secondary rhinaria distributed III 0-6, IV 0-2, and alatae have them distributed III 5-7, IV 2-3. On stems and flowers of Carthamus tinctorius and other Carthamus spp., attended by ants. In north Africa, Lebanon, Iran, Kazakhstan, Pakistan and north-west India (Naumann-Etienne & Remaudière 1995). Kadyrbekov (2005b) records it also from several other composite genera. Specimens in the BMNH collection from Atractylis flava and Hedypnois rhagadioloides in Egypt and from Cirsium lanceolatum in North Yemen may also be this species, although these are intermediate in some respects between carthami and anuraphoides. North African records of anuraphoides and pseudocardui from Carthamus are possibly referable to carthami. P. echinopis on Echinops is very similar and possibly a synonym.
Protaphis chondrillae (Mordvilko in Tarbinsky and Plaviltchikov) Apterae are dark green with slight grey wax film, with black siphunculi and dark brown dorsal markings; BL 1.8-1.9 mm. On roots or subterranean stems of Chondrilla sp., ant-attended. Southern Kazakhstan (Kadyrbekov 2001b).
Protaphis cousiniae Kadyrbekov Apterae are dark greenish with slight grey film; BL 1.6-2.3 mm. On roots of Cousinia spp., attended by ants (Kadyrbekov 2001b, 2005d, 2017a). In Central Asia (south and east Kazakhstan). Very similar to P. terricola.
Protaphis debilicornis (Gillette & Palmer) Apterae are dark olive buff; BL 1.6-2.1 mm. In curled leaves and on stems of Helianthus spp., widely distributed in USA. Monoecious holocyclic, with oviparae and alate males in September (in Colorado; Palmer 1952). Lagos et al. (2014) transferred this species to Protaphis.
Protaphis delottoi Eastop Colour of apterae in life unknown, probably dark green or brown dusted with wax; BL 1.5-1.8 mm. Apterae have 0-4 secondary rhinaria on ANT III, and alatae have them distributed III 9-13, IV 0-4. On Pluchea dioscoridis, probably at base of plant with ants. In Eritrea, and possibly Nigeria (Eastop 1961a).
Protaphis dudichi (Börner) Colour of apterae in life unrecorded; BL 1.5-2.0 mm. On roots of Matricaria and Tripleurospermum spp.in central and eastern Europe, and also in western Siberia (Stekolshchikov et al. 2007a). Börner’s material was redescribed by Tashev (1964a), who gave some possible differences from the very similar P. anthemidis, but these could be host-related, and it has been treated in catalogues as a synonym. However, it resembles P. carlinae even more closely, and further work is needed to establish whether one, two or three species are involved.
Protaphis echinaceae (Lagos & Voegtlin) Colour of apterae in life is unrecorded; BL 1.5-2.0 mm. On underside of flowerhead and leaves of Echinacea angustifolia, attended by ants, which often build shelters over leaf-feeding colonies (Lagos & Voegtlin 2009). Only known from Minnesota, USA. Alatae have secondary rhinaria distributed ANT III 2-15, IV 3-10, V 1-5. Sexual morphs and life cycle are unknown. Lagos et al. (2014) transferred this species to Protaphis.
Protaphis echinopis (Hille Ris Lambers) Colour of apterae in life is unrecorded, probably dark green with some wax; BL 1.2-2.2 mm. Apterae without secondary rhinaria, alatae with them distributed III 6-10, IV 0-1. On stems and leaves of Echinops spinosus, and possibly other Echinops spp., in north-east Africa (Egypt, Eritrea, Ethiopia) and Middle East (Israel, Yemen). Records of P. anuraphoides and P. pseudocardui on Echinops in this region should perhaps all be referred to this species. However, the distinction from P. carthami on Carthamus is unclear, and P. echinopsicola (Kadyrbekov), described from Echinops albicaulis in Central Asia, seems to be intermediate in morphology between A. echinopis and A. terricola.
Protaphis elatior (Nevsky) Apterae are dark green, with a fine blue pruinose covering, black eyes (cf. elongata) and black transverse bands on posterior abdomen; BL 1.5-2.0 mm. Described from roots of Artemisia annua, and subsequently recorded from other Artemisia and Seriphidium spp. Records from various other Compositae/Asteraceae (Ivanoskaya 1960; Narzikulov et al., 1971) are perhaps referable to other Protaphis spp. Ant-attended. In Russia, Iran (Rezwani et al. 1994) and Central Asia (Kadyrbekov 2001b).
Protaphis elongata (Nevsky) Apterae are glaucous green with blue-grey pruinose covering, dark head and black markings on thorax. Eyes are red (cf. elatior). BL 1.4-1.7 mm. On roots and subterranean stems of Artemisia and Seriphidium spp., attended by ants. There are also a few records from other Compositae/Asteraceae, e.g. Xanthium strumarium. In Uzbekistan, south-east Kazakhstan, Ukraine, and Iran (BMNH collection, leg. R. van den Bosch), and also reported from Poland, Rumania and Moldova. Some records of this species including those from western Siberia and Denmark are possibly ascribable to P. miranda (Kadyrbekov 2001b).
Protaphis evansi (Eastop) (Fig.57a) Apterae are dull, mottled greenish or brownish red; BL 1.2-1.9 mm. On Vernonia glabra, at base of stem with ants. Also recorded from Leucospermum cordifolium and Hypochaeris radiata. In southern Africa (Botswana, Tanzania, Zimbabwe, South Africa). Kadyrbekov (2001c) placed this species in a new genus (Eastopiella), but some of the characters that he described (shape of cauda, presence of median ocellus in aptera, number of accessory hairs on R IV+V) do not agree with type material in the BMNH collection, and in most respects P. evansi seems to be a typical Protaphis.
Protaphis filaginea (del Guercio) Apterae are dark green to brownish; BL not recorded. Described from Filago germanica (= F. vulgaris), and also possibly on Filago (=Logfia) gallica, if it is the same species as Aphis filaginis Lichtenstein, which is a nomen nudum. In southern France and Portugal.
Protaphis formosana (Takahashi) Apterae are dark green, with black head, antennae, siphunculi and cauda; BL c.1.8 mm. On Blumea spp.in Taiwan. Apterae have 1-5 secondary rhinaria on ANT III, alatae have 6-8 on III, and 0-1 on IV.
Protaphis hirsuta (Nevsky) Apterae (immature?) are dark green to greenish brown, wax-coated; BL c.1.2 mm. On undersides of leaves and stems of an Artemisia sp. (?vulgaris) in Uzbekistan. Described in Cryptosiphum, and subsequently placed in Xerobion, but apparently the type specimens are an immature Protaphis (Kadyrbekov 2014b).
Protaphis iliensis Kadyrbekov Apterae are dark greenish with grey wax film, with dark dorsal sclerotisation, siphunculi and cauda; BL 1..6-2.3 mm. On stems and flowers of Acroptilon australe (= Rhaponticum repens), attended by ants; subsequently recorded from Karelinia caspia (Kadyrbekov & Aoitzhanova 2005). Sedighi et al. (2020b) also record it from Artemisia (= Seriphidium) sieberi and Gundelia sp.,and provide a redescription. In Kazakhstan and Iran..
Protaphis kareliniae Kadyrbekov, Renxin & Shao Apterae are yellow-green, with slight grey wax film and dark brown head, siphunculi and dorsal markings; BL 2.1-2.1 mm. Secondary rhinaria in alata (1 specimen) distributed III 6-8, IV 1. On roots of Karelinia caspia, attended by ants. In Xinjiang-Uygur region, western China (Kadyrbekov et al. 2002).
Protaphis knowltoni (Hottes & Frison) Apterae are greenish blue to yellowish green, with black dorsal markings and greyish pruinose reticulation; BL 1.8-2.3 mm. Apterae have secondary rhinaria distributed III 0-13, IV 1-8, V 0-3, VI BASE 0-1; alatae have them distributed III 20-32, IV 7-15, V 3-8. In ant-attended colonies on roots of Taraxacum officinale and also recorded from Achillea millefolium (Palmer 1952) and Dahlia sp. (BNHM collection, leg. A.G. Robinson). Palmer also records it from Zea mays, but this needs confirmation as there is possible confusion with P. middletonii. In USA and Canada (Manitoba). Monoecious holocyclic, with oviparae in September (males undescribed). 2n=8.
Protaphis kvavadzei Bardjadze & Özdemir Colour of apterae in life unknown; BL 1.5-1.9 mm. Apterae have secondary rhinaria distributed III 0-6, IV 0-3, V 0; alatae (2 specimens) have them distributed III 12-19, IV 2-3, V 0. On Eryngium campestre in central Turkey (Barjadze et al. 2014).
Protaphis lactucicola Kadyrbekov Apterae are dark greenish with slight grey wax film, brown head, usually a brown central dorsal transverse mark just anterior to siphunculi, and dark brown siphunculi; BL 1.6-1.9 mm. In ant-attended colonies on roots of Lactuca and Crepidiastrum in south-east Kazakhstan. Very similar to P. pseudocardui (q.v.).
Protaphis middletonii (Thomas) (= Aphis armoraciae Cowen, = Aphis maidiradicis Forbes, = Aphis menthaeradicis Cowen) Plate 7e Apterae are pale green, bluish green, grey-green or olive-green, often dusted with greyish wax, and with variably-developed dark dorsal markings or cross-bands, more apparent in apterae than in alatae (see influentialpoints.com/Gallery); BL 1.5-2.5 mm. Apterae have secondary rhinaria distributed III 0-17, IV 0-8, V 0-5*, and alatae have them distributed III 4-22, IV 0-9, V 0-6*. Ant-attended colonies occur commonly in western and central USA on roots of plants in numerous families including Compositae/Asteraceae, Brassicaceae, Poacaeae, Lamiaceae and Apiaceae. Aphids of this group are also recorded from Compositae/Asteraceae in Brazil, and from Opuntia roots in Australia and South Africa. Records from Centaurea in Turkey (Akyürek et al. 2011) and from Taraxacum in Algeria (Laamari et al. 2013) need additional confirmation. Eastop & Blackman (2005) concluded on the basis of multiple discriminant analysis that a single name should be applied to this group, the oldest available being middletonii Thomas. Monoecious holocyclic, but anholocyclic overwintering is probably also common. In the North American literature, populations identified as armoraciae are stated to produce alate males, whereas maidiradicis and menthaeradicis have apterous males, and oviparae of menthaeradicis are said to have very few scent glands on their hind tibiae (Palmer 1952). However, the criteria for naming these populations are unclear, and experimental work is needed to show whether these differences are of any taxonomic significance. Two other North American taxa described by Pack & Knowlton (1929), Aphis crypta and A. gutierrezis (as Anuraphis gutierrezis),are closely related and probably part of this group. 2n=8.
*the numbers of rhinaria given in Blackman & Eastop (2006) were erroneous
Protaphis miranda Kadyrbekov Apterae are dark greenish with slight grey film, dark reddish eyes, black siphunculi and green cauda: BL 1.4-1.8 mm. In ant-attended colonies on roots of Artemisia and Seriphidium spp. in Kazakhstan (original description), west Siberia (Ivanoskaya, 1977, as alexandrae) and Europe (Denmark, Poland – Heie, 1986, as elongata; Czech Republic – BMNH collection, leg. J. Holman), and also recorded from two Tanacetum spp. (Kadyrbekov 2017a). Probably monoecious holocyclic; an ovipara from A. campestris in Poland (BMNH collection, leg. L. Olesinski) may be this species.
Protaphis pseudocardui Theobald (Fig.31c) Apterae are dark green or brown with covering of grey mealy wax, short black siphunculi, and often there are a pair of dark spots on dorsal abdomen just anterior to siphunculi; BL 1.5-2.0 mm. In ant-attended, often dense colonies on stems, upper sides of leaves, flowerheads, or on root collars. Described from “thistles (Carduus sp.)” in South Africa; however this could have been Sonchus,which is widely known as “thistle” in Africa. Since recorded from other parts of Africa, Mediterranean region, the Middle East and Georgia (Barjadze 2008), and from various other Compositae (Arctotheca, Berkheya, Carduus, Carthamus, Cichorium, Hypochaeris, Lactuca, Pluchea, Pulicaria, Vernonia), although some of these records may be referable to other closely-related species, including the terricola group. 2n=8.
Protaphis rutae (Nevsky) Apterae are dark green, with a light wax covering, siphunculi black and cauda green; BL 1.4-2.0 mm. On Ruta sieversii (= Haplophyllum acutifolium?), forming large colonies on upper and undersides of leaves. In Central Asia (Uzbekistan). This species was redescribed and transferred to Protaphis by Kadyrbekov (2001b).
Protaphis scorzonerae (Mordvilko) Apterae are brownish green, covered with “rather bright azure-blue velvet-like powder” (Ghilarov 1937, as Xerophilaphis scorzonerae); BL 1.7-2.3 mm. In ant-attended colonies on roots (where they may feed in lesions produced by the ants; Ghilarov1937) and at bases of leaves, of Compositae/Asteraceae (Taraxacum, Hieracium, Scorzonera, Sonchus). Austria (BMNH collection, leg. H. Franz), central Russia, Ukraine, north Caucasus, and eastward to Kazakhstan.
Protaphis sonchi (Narzikulov & Umarov) Apterae are dark green, wax-dusted; BL c. 1.6 mm. Alatae have secondary rhinaria distributed III 10-11, IV 3-4, V 3. On stems and leaves of Sonchus asper in Afghanistan, and subsequently recorded from S. oleraceus in Kazakhstan (Kadyrbekov & Aoitzhanova 2005). Not clearly differentiated from other members of the terricola group (q.v.).
Protaphis terricola Rondani Plate 7f Apterae are dark green, with pruinose covering; BL 1.3-2.0 mm. Apterae have secondary rhinaria distributed III 0-12, IV 0-4, alatae have them distributed III 7-13, IV 1-4. On Centaurea spp., and also recorded from plants in several other composite genera (see below). On ground rosettes of young plants, later on stems and inflorescences. In southern, central and eastern Europe (France, Austria, Switzerland, Italy, Spain, Portugal, Canary Islands, Germany, Poland, Czech Republic, Hungary, Bulgaria, Ukraine), Iran (Rezwani 1990), Turkey, Israel, Kazakhstan (Lagos et al. 2014), Pakistan (Naumann-Etienne & Remaudière 1995), Egypt, Sudan, and introduced to South America (Argentina, Brazil, Chile). Monoecious holocyclic (Börner 1952, as alexandrae).
[There are a number of Protaphis species described from Compositae/Asteraceae in Europe, Central Asia and north Africa that cannot be satisfactorily distinguished from terricola. These include alexandrae Nevsky, hartigi Hille Ris Lambers, hyaleae Kadyrbekov and centaurea Gomez-Menor, all described from Centaurea and related genera; kenanae Müller & Tigani from Sonchus cornutus;and ignatii Gomez-Menor from an unidentified composite plant (but later recorded from Carlina sp.). García Prieto et al. (2004) synonymised centaurea with terricola. Hille Ris Lambers (1967) compared some of these species with terricola. Kadyrbekov (2001b) described several more species in this group (ancathiae, aralensis, cousiniae, turanica) and provided a key to central Asian Protaphis, but did not compare any of them with terricola. It needs to be demonstrated, and confirmed by experimental studies, that the differences are more than the result of local or environmental variation.] 2n=8.
Protaphis tetradymia Knowlton Apterae are bluish green with blackish antennae, siphunculi and cauda; BL 1.3-1.8 mm. Alatae are undescribed. On aerial parts of Tetradymia canescens in Idaho and Utah, USA. Monoecious holocyclic with oviparae in September (BMNH collection, leg. G.F. Knowlton).
Protaphis turanica Kadyrbekov Colour of apterae in life unrecorded, probably dark green; BL 1.5-2.1 mm. On roots of Cirsium spp.in Central Asia (southern Kazakhstan; Kadyrbekov 2001b). Very similar to P. terricola, and possibly a synonym.
|
Protohormaphis Shaposhnikov & Gabrid |
Hormaphidinae: Hormaphidini? |
One conifer-feeding species of uncertain phylogenetic position. A cladistic study by Chen et al. (2014), suggests that it may occupy a basal position in the Hormaphidini, along with Tsugaphis. Only the apterous vivipara is known.
Protohormaphis piceae Shaposhnikov & Gabrid (fig. 65A) Apterae are oval, slightly flattened, dark green; BL c. 1.0 mm. Immatures are lighter bluish green with white wax markings. Found feeding on needles of Picea pungens in May-June, in Kirgizia (Shaposhnikov & Gabrid 1987). The life cycle is unknown.
|
Protopterocallis Richards |
Calaphidinae: Panaphidini |
Four eastern North American species on Carya related to Monelliopsis, but larger and more robust, and with a shorter antennal terminal process. All adult viviparae are alate. Bissell (1978) reviewed the genus. All stages are said by Bissell to feed on the upper surface of the leaves, usually along mid-ribs of basal leaflets. Quednau (2003) provided a key and illustrations of all available morphs.
Protopterocallis fumipennella (Fitch) (= P. canadensis Richards) Alatae are broad-bodied, with abdomen yellow-orange, yellowish green or olive-green dusted with wax, prothorax darker, dorsal abdominal tubercles brown-tipped; BL 2.0-2.2 mm. They feed singly or in small colonies on undersides(?) of leaflets of Carya spp., especially cordiformis (but not illinoensis), in eastern USA and Canada (Quednau 1966, Bissell 1986). Oviparae and males occur in September-October (Bissell 1978).
Protopterocallis gigantea Bissell (fig. 33L,M) Alatae are broad-bodied, with abdomen dull yellow with irregular streaks of orange or red, dorsal abdominal tubercles brown-tipped and forewing veins (especially Cu1b) dark brown (see influentialpoints.com/Gallery); BL 2.5-3.0 mm. On Carya spp. (but not illinoensis) in eastern USA (D.C., Maryland, Georgia). Oviparae and males occur in October (Bissell 1978). 2n=10.
Protopterocallis pergandei Bissell (fig. 33K) Appearance in life is unknown; alata are broad-bodied, BL 2.5-2.9 mm. Described from Carya sp. in Florida, USA. An ovipara was collected on Carya pallida in Georgia in November (Bissell 1978).
Protopterocallis quadrata Bissell (fig. 33J) Alata are broad-bodied, yellowish; BL 1.7-2.8 mm (Bissell 1978). On Carya tomentosa and Carya sp. in eastern USA (Pennsylvania, D.C., Georgia). Biology and sexuales are not recorded.
|
Protrama Baker |
Lachninae: Tramini |
Five palaearctic species with greatly elongate hind tarsi and siphunculi present as pores on pigmented flat cones. Apterae are often alatiform with variable dorsal sclerotisation. Like Trama they mostly live on roots of Compositae, and are mainly anholocyclic, at least in western Europe. Accounts are available for western Europe (Eastop 1953, Heie 1995), the UK (Blackman et al. 2019b), the Iberian peninsula (Nieto Nafría et al., 2002a), Poland (Szelegiewicz 1978), Central Asia (Narzikulov 1963) and India (A.K. Ghosh 1982b).
Protrama flavescens (Koch) Apterae (all alatiform) are dirty or creamy white, greyish or greenish with variable dark dorsal cross-bars; BL 2.7-3.8 mm. Alatae and intermediates between alatae and apterae are very common. On roots of Artemisia spp., and sometimes on Achillea, Chrysanthemum and certain other Compositae/Asteraceae, mostly in tribe Anthemideae. In Europe, Iran (Hodjat 1993), and perhaps in India if P. longitarsus sclerodensus is a synonym. Anholocyclic. 2n=40-42.
Protrama longitarsus (Ferrari) Appearance in life is unknown, probably rather green or grey with darker dorsal cross bands; BL 2.8-3.8 mm. On roots of Artemisia campestris in Italy and Switzerland (Eastop 1953), and also recorded from Germany, Poland and Czech Republic. Apterae and alatae collected on A. gmelinii in northern Japan (BMNH collection, leg. V.F. Eastop) may also be this species. Börner (1952) recorded it from Doronicum caucasicum in Germany.The life cycle is unknown; it is probably mainly anholocyclic, but oviparae were collected in October in Germany (BMNH collection, leg. H Zwölfer). No satisfactory morphological discriminants to distinguish P. longitarsus from P. ranunculi have been found.Kumar (1973) described heavily pigmented specimens without secondary rhinaria on ANT III in India as a subspecies, P. longitarsus ssp. sclerodensis Kumar, and similar aphids have since been recorded from roots of various plants in India (A.K. Ghosh 1982) and Pakistan (Naumann-Etienne & Remaudière 1995). However, specimens under the name sclerodensis in the BMNH collection seem indistinguishable from P. flavescens.
Protrama radicis (Kaltenbach) Plate 3f Apterae are dirty white to pale yellow or pale brownish green, with brown siphuncular cones (see influentialpoints.com/Gallery) ; BL 2.5-3.4 mm. Almost all apterae are to some extent alatiform. True alatae have been recorded (see influentialpoints.com/Gallery). In ant-attended colonies on roots of various Compositae-Cynareae (Arctium, Carduus, Centaurea, Cirsium, Cynara). However Eastop (1953) found that specimens collected from Cynara refused to feed on Cirsium or Centaurea, and those from Cirsium could not be transferred to Arctium or Centaurea, suggesting the possible existence of host races. In Europe (not Iberian peninsula), south-west and central Asia (Kadyrbekov, 2005d). A form on Centaurea iberica in Tajikistan was described as a subspecies, P. radicis asiatica Narzikulov. Apparently completely anholocyclic. 2n=c.60.
Protrama ranunculi (del Guercio) Apterae (all alatiform) are dirty or creamy white, greyish or greenish with variable dark dorsal cross-bars; BL 2.8-3.7 mm. On roots of Ranunculus spp. attended by ants. In Europe, west Siberia, and Central Asia. 2n=c.36.
Protrama taraxaci Kan Apterae are brownish green; BL c.2.5 mm. On roots of Taraxacum officinale in Uzbekistan.
|
Pseudacaudella Börner |
Aphidinae: Macrosiphini |
One moss-feeding species similar to Myzodium but with lower and less scabrous antennal tubercles, and adaptations for surviving adverse conditions without a sexual phase.
Pseudacaudella rubida Börner Plate 16h (Fig.44d,f) Apterae are shining olive green to brown with rusty patches at bases of siphunculi; BL 0.7-1.0 mm. Alatae have secondary rhinaria distributed III 7-15, IV 3-7, V 0-2. On mosses in various genera; Acrocladium, Climacium, Dicranum, Hylocomium, Mnium, Pleurozium, Polytrichum, Pseudoscleropodium, Thuidium). Europe, Kazakhstan (Kadyrbekov 2017a), Morocco (BMNH collection, leg. P.N. Lawrence), Brazil (leg. S. Lazzari) and USA (North Carolina), Panama (Quiros et al. 2009), and alatae have been trapped in New Zealand (leg. J.A. & P.A. Ware). It seems able to survive in dry as well as humid habitats, and is adapted for anholocycly, with a specialised overwintering second instar having a dark sclerotic cuticle and a wax coat (Müller 1973b, Tinguely 1993). However there is possibly host alternation from Sorbus in Ukraine (as Staegeriella sp.; Bozhko, 1976:30).
|
Pseudaphis Hille Ris Lambers |
Aphidinae: Macrosiphini |
Three species in Africa and Asia, related to Sitobion but differing by their completely undeveloped antennal tubercles and several other characters. The siphunculi only have a small amount of subapical reticulation and this varies from 1-5 rows of polygonal cells. Aldryhim & Ilharco (1997) revised the genus.
Pseudaphis abyssinica Hille Ris Lambers Apterae are pale green with variably developed dark dorsal cross-bands; BL 1.4-2.2 mm. In ant-attended colonies on Rosa spp. and Rubus apetalus in east Africa (Eritrea, Ethiopia, Burundi), and also found on unidentified Poaceae in Yemen (Aldryhim & Ilharco 1997), indicating that host alternation probably occurs. Alatae have been reported from India (A.K. Ghosh et al.1971f).
Pseudaphis arabica Aldryhim & Ilharco Apterae are brown with darker head, thorax and dorsal abdominal cross-bands; BL 1.7-2.2 mm. In flower-heads of Pennisetum setaceum in Saudi Arabia and Eritrea (Aldryhim & Ilharco 1997). Closely related to P. sijui, differing only in its larger size and more extensive sclerotisation, which differences could be environmentally determined.
Pseudaphis sijui (Eastop) Plate 26g Apterae are dark olive-green to brown, slightly wax-dusted; BL 1.1-1.6 mm. In flower-heads of various grasses (Cenchrus, Exotheca, Hyparrhenia, Miscanthus, Pennisetum, Setaria, Sorghum), and also Cyperus sp., in east Africa (Kenya, Burundi). Remaudière & Autrique (1985) give further information.
|
Pseudasiphonaphis Robinson |
Aphidinae: Aphidini |
One North American species with very small flangeless siphunculi, well-developed lateral tubercles and extensive wax secretion (Robinson 1965a).
Pseudasiphonaphis corni (Tissot) Spring colonies, on undersides of leaves and along young shoots of Cornus microcarpa in Florida, contained brown apterae (BL 1.7-2.1 mm) underneath an extensive light greyish white woolly wax secretion.  (see influentialpoints.com/Gallery); Alatae have dark wing veins and a reddish brown abdomen (Tissot 1929). Further north in the USA (Pennsylvania, Illinois, New York), there are very few records from Cornus, and dense colonies of this species occur most commonly on main stems and flowerstalks of Pilea pumila (Urticaceae), again obscured by copious flocculent wax. A colony has also been found on a labiate, Scutellaria laterifolia (Robinson 1965a). Alate males have been found in October on Pilea (Hottes & Frison 1931, as Asiphonaphis anogis). Host alternation is indicated, but requires confirmation; there are no records of the return migration to Cornus. In eastern USA and Canada. In Florida, anholocyclic populations persist all year on Cornus (Tissot 1929).
|
Pseudessigella Hille Ris Lambers |
Lachninae: Cinarini |
One pine-feeding species in Pakistan and India related to Eulachnus and Essigella; Sorensen (1990, 1991) discussed its relationship to these other genera. Molecular data have now placed it as a sister-group to Essigella (Théry et al. 2018b).
Pseudessigella brachychaeta Hille Ris Lambers Apterae are pale green, narrowly elongate; BL 2.0-2.6 mm. On needles of Pinus griffithii (= wallichiana) in Pakistan (Hille Ris Lambers 1966c) and northern India. Apterae and oviparae have also been collected on the same host in India (Jammu and Kasmir; Kanturski et al. 2016). Alatae are unknown. A record from Turkey (Görür et al. 2011a) requires confirmation. Kanturski et al. (2016a) made SEM studies of its morphology and the ultrastructure of antennal and labial sensilla, and Kanturski et al. (2017a) described the ovipara and dwarfish male collected in late September-October.
|
Pseudobrevicoryne Heinze |
Aphidinae: Macrosiphini |
Three palaearctic species related to Brevicoryne but with very short siphunculi, first tarsal segments with 3,3,2 hairs, and apterae with secondary rhinaria. A fourth species, Siphonatrophia (Lacusaphis) aetherelaca, described fron an unidentified crucifer in China (L. Zhang & G. Zhang 2002a), also belongs in this genus.They roll the leaves of Brassicaceae.
Pseudobrevicoryne buhri Börner Apterae are yellowish to olive-brown, powdered with grey wax; BL 1.5-2.3 mm. Alatae have secondary rhinaria distributed III 35-56, IV 5-11, V 3-6. In colonies on upper sides of leaves of Barbarea spp., which are rolled longitudinally upwards. Also recorded from Brassica campestris (Shaposhnikov 1964). In northern Europe, including UK, Scandinavia, northern Germany and Poland, and south-eastward to Carpathians and Ukraine. Monoecious holocyclic; fundatrices have been found in mid-May in southern England, but sexual morphs have not been described.
Pseudobrevicoryne erysimi Holman Apterae are greyish green to pale green, covered in wax powder, with dark brown to black appendages; BL 1.1-1.8 mm. Alatae have secondary rhinaria distributed III 20-31, IV 2-5, V 0-1. Living on upper sides of leaves of Erysimum spp., which are rolled upwards into compact tubes and usually become reddish to dirty purple. Known from Czech Republic, Hungary and Kazakhstan. Monoecious holocyclic, with apterous males (original description).
Pseudobrevicoryne leclanti Petrović & Remaudière Apterae are green, covered in white wax powder; BL 1.3-1.9 mm. Alatae have secondary rhinaria distributed III 30-38, IV 4-7, V 1-3. On upper sides of leaves of Arabis alpina, causing lateral parts of leaves to fold upwards towards midrib. Known only from Montenegro (former Yugoslavia). Monoecious holocyclic with apterous males and oviparae in mid-August (Petrović & Remaudière 2002).
|
Pseudocercidis Richards |
Aphidinae: Macrosiphini |
Two species in western North America, possibly related to Sitobion but with first tarsal segments bearing 5 or 6 hairs, short cylindrical siphunculi, and a short cauda.
Pseudocercidis rosae Richards Plate 26h Apterae are covered with white wax powder; BL 2.2-3 mm. They feed inside curled of folded leaflets of terminal leaves of Rosa spp., usually in shaded conditions, in western USA and Canada. Monoecious holocyclic, although there is an unexplained collection from Lupinus nootkatensis in Alaska (BMNH collection, leg. F.C. Hottes). Alatae are rare. Oviparae and alate males occur in September (Robinson 1963). 2n=12.
Pseudocercidis tutigulus (Hottes) Apterae are pale yellow-green; 2.3-2.7 mm. On undersides of leaves and flower stems of a Corydalis spp. in western USA (Colorado, Utah, Idaho, Washington) and British Columbia (A. Jensen, aphidtrek.org). Heteroecious holocyclic, with a sexual phase on wild Rosa, on whichalate males and oviparae were found in September (Hille Ris Lambers 1961, as Adactynus tutigula). [This species has previously been placed in Kakimia (Palmer 1952) and Acyrthosiphon (e.g. Blackman & Eastop 2000), but was transferred to Pseudocercidis by Eastop & Blackman (2005).]
|
Pseudochromaphis Zhang |
Calaphidinae: Pseudochromaphidini |
One east Asian species perhaps related to Sinochaitophorus. All viviparae are alatae with characteristically marked maculate forewings lacking a radial sector.
Pseudochromaphis coreana (Paik) (fig. 51A) The general colour is not recorded, but the black head and prothorax with pale patches, the maculate forewings and the pattern of small dark spots on the dorsal abdomen are probably very distinctive in life; BL of alata 1.1-1.6 mm. On Hemiptelea davidii in China (Zhang & Zhong 1982c) and Korea (Paik 1965); subsequently also recorded from Korea on Ulmus pumila and Ulmus sp. (Quednau & Lee 2001). An ovipara collected in October is illustrated by Quednau (2003).
|
Pseudoepameibaphis Gillette & Palmer |
Aphidinae: Macrosiphini |
Four North American species on Artemisia, with the typical stilletto-shaped R IV+V of aphids feeding on Anthemideae, and related to Epameibaphis, Flabellomicrosiphum and the palaearctic genus Coloradoa,but with characteristically-shaped siphunculi. Palmer (1952) reviewed the genus.
Pseudoepameibaphis essigi Knowlton & Smith Apterae are whitish yellow to orange-green, with pale appendages; BL 0.7-1.1 mm. On Artemisia tridentata in western USA. Monoecious holocyclic; oviparae and alate males found in Utah in October (BMNH collection, leg, G.F. Knowlton).
Pseudoepameibaphis glauca Gillette & Palmer Apterae are glaucous green, pulverulent, with pale appendages; BL 1.2-1.5 mm. On leaves and stems of Artemisia frigida in western USA. Monoecious holocyclic with apterous males (Palmer 1952). The account by Knowlton & Smith (1938a) under this name applies to P. tridentatae.
Pseudoepameibaphis tridentatae (Wilson) Plate 15f (Fig.12g) Apterae are pale yellowish with frosted appearance due to numerous pale fan-shaped dorsal hairs; BL 1.3-1.5 mm. On leaves and stems of Artemisia tridentata in western USA, and collected from Artemisia cana in Saskatchewan, Canada (BMNH collection, leg. W.R. Richards). Monoecious holocyclic; oviparae and alate males were collected in Utah in October-November (BMNH collection, leg. G.F. Knowlton).
Pseudoepameibaphis xenotrichis Knowlton & Smith Apterae are greenish, with whitish appearance due to numerous pale fan-shaped dorsal hairs; BL 0.9-1.2 mm. On Artemisia tridentata in western USA.
|
Pseudomegoura Shinji |
Aphidinae:Macrosiphini |
Two oriental species related to Aulacorthum but with a black head, distinctly clavate siphunculi and a dark cauda with only five hairs (W. Lee et al. 2011a, as Neoaulacorthum, which is an objective synonym of Pseudomegoura Shinji 1922; see Nieto Nafría & Favret 2014)).
Pseudomegoura magnoliae (Essig & Kuwana) Plate 22h Apterae are distinctly coloured with black front of head and reddish prothorax, rest of body yellow-green to green; femora mostly black, tibiae and antennae usually wholly black, siphunculi pale basally and dark distally, cauda dark; BL 2.3-3.9 mm. Polyphagous, feeding mainly on leaves of plants in over 20 different families, including Citrus and many ornamental shrubs and trees. Mainly anholocyclic, in Japan, China, Korea and India (Blackman & Eastop 2000, p.244, as Aulacorthum); with a “relict” holocycle on Sambucus in Japan. Matsuka & Imanishi (1982) studied its life cycle on Sambucus sieboldiana near Tokyo, where populations overwinter both as viviparae and, less commonly, as eggs. Clones descended from fundatrices produced males and viviparous females, but very few oviparae, so the sexual phase was almost non-existent in that population. Males are also recorded from India (Raychaudhuri et al. 1980b). Indian records are mostly from Cucurbitaceae and need confirmation due to confusion with Aulacorthum spinacaudatum (q.v.). A record from Turkey (Kaygin et al. 2009) also requires further confirmation. 2n=12 (ex. cult. on potato, Japan).
Pseudomegoura nipponica (Essig & Kuwana) Apterae are yellowish red to pink with black head, siphunculi and cauda; BL 2.2-2.5 mm. On leaves and stems of Paederia foetida in east Asia (Japan, Taiwan, China, Thailand, Korea, Java, north-east India), but there are Hong Kong records from Gymnema tingens and Mikania micrantha (the latter a large colony; Martin & Lau 2011), indicating polyphagous tendencies. Anholocyclic in Taiwan, monoecious holocyclic near Tokyo with oviparae and alate males in late October (Takahashi 1923, as Macrosiphum paederiae).
|
Pseudopterocomma MacGillivray |
Chaitophorinae |
Two poplar-feeding species only known from Canada and north-eastern USA, related to Chaitophorus but with a distinctive, long, thick ANT PT clothed with hairs like the rest of the antenna, and pore-like siphunculi. Richards (1972a) provided a key.
Pseudopterocomma canadensis Richards Apterae are grey or black; BL 3.0-3.5 mm. Alatae have dark-bordered wing veins. On Populus tremuloides in Canada (Manitoba, Ontario, Saskatchewan). The feeding position on the plant is not recorded, but is likely to be low down on the stem or suckers, or possibly below ground level. Sexual morphs and life cycle are unknown.
Pseudopteocomma hughi (MacGillivray) Apterae are cream-coloured; BL 2.3-3.0 mm. Alatae have dark-bordered wing veins. On roots of Populus tremuloides, attended by ants. In Maine, USA (BMNH collection, leg. G. Remaudière) and Canada (Manitoba, New Brunswick, Quebec). Fundatrices occur in May, and apterous males and oviparae in September (MacGillivray 1963b).
|
Pseudoregma Doncaster |
Hormaphidinae: Cerataphidini |
About 12 species of wax-dusted aphids living mostly on either bamboos or Zingiberidaceae, with a few species on other Poaceae and some other monocotyledons in south-east Asia. Most of the species were originally described in Oregma, and may be found in accounts of Oregma until 1966. The long-suspected host alternation to Styrax was confirmed for two species in Taiwan (Aoki & Kurosu 1991). There is a remarkable polymorphism in the first or second instar, some nymphs being pseudoscorpion-like and functioning as soldiers for the colony (Aoki 1987). Accounts are available for Java (Noordam 1991), India (A.K. Ghosh 1988) and China (Qiao et al. 2018). Aoki et al. (2007) provided a key to bamboo-feeding species. Fukatsu et al. (1994) and Stern (1998) considered the relationship with other Cerataphidini and the evolution of the soldier caste. Phylogenetic relationships of Pseudoregma and Ceratovacuna based on two mitochondrial and two nuclear genes were studied by R. Zhang et al. (2013). Aoki & Kurosu (2010) reviewed life cycles and biology.
Pseudoregma alexanderi (Takahashi) Apterae are rather elongate pear-shaped, pale brown to brownish black with a purple tinge, lightly dusted with wax powder that is often thicker on the posterior abdomen; BL 2.2-4.3 mm. Large colonies attack the young shoots and leaves of bamboos such as Dendrocalamus, and there are also records from Miscanthus and Echinochloa. In China, Taiwan, Sumatra, Nepal and north-east India. Soldier behaviour was studied by Aoki et al. (1981). The life cycle is uncertain; alate sexuparae are produced in May in Taiwan, but the primary host is unidentified and populations also persist throughout the year on Dendrocalamus (Aoki et al. 1981). 2n=12.
Pseudoregma baenzigeri Aoki, Kurosu & Sirikajornjaru Apterae are greyish green; BL 2.8-3.6 mm. Forming large dense colonies on Dendrocalamus sp. in northern Thailand (Aoki et al. 2007). Many first instar pseudoscorpion-like soldiers were present in the colonies, and alate sexuparae were produced from late September to mid October. Predation by two syrphid species, by pyralid larvae (Dipha aphidivora), and possibly by rodents, was recorded.
Pseudoregma bambucicola (Takahashi) Galls on Styrax suberifolia in Taiwan consist of clusters of elongate pod-shaped sub-galls with petiolate bases (cf. P. koshunensis) and apical openings, arising from a single point on the stem (Aoki & Kurosu 1992). Gall populations include second instar soldier larvae. Alatae emerge from galls in July-September to colonise bamboos, and alate sexuparae return in December. Apterae on bamboos in India identified as this species are in life dark olive green to grey, with white wax dusted over dorsal surface and concentrated in a pair of tufts on posterior abdomen; BL c.3mm (see aphids of Karnataka website and influentialpoints.com/Gallery – although the identity of the Indian populations needs to be confirmed, see below). Preserved specimens are apparently almost indistinguishable from those of P. carolinensis (q.v.). P. albostriata Liao 1976, described from Bambusa oldhami and Dendrocalamus giganteus in Taiwan, is regarded as a synonym of P. bambucicola (Remaudière & Remaudière 1997), but that species has four longitudinal wax stripes in life, not three as in P. carolinensis. P. cantonensis Takahashi, differentiated by having greater sclerotisation of the dorsum, is another possible synonym within the bambucicola/carolinensis group.
Dense, ant-attended colonies occur on the shoots, young stems and leaf bases of Bambusa spp., especially B. multiplex. First instar soldiers occur within the colonies. Sunose et al. (1991) studied population dynamics of P. bambucicola on B. multiplex in Japan, Arakaki & Yoshiyasu (1988) studied predation by a pyralid moth, and Ôhara (1985a,b) and Al Siddiqui et al. (2019) studied the defensive behaviour of soldiers. In Japan, parthenogenetic reproduction continues throughout the year; sexuparae are produced in autumn (Sunose et al. 1991), although the primary host is not available. The role of soldiers in defending sexuparae on bamboos was discussed by Sunose et al. (1991), Îto et al. (1995) and Aoki (1997). The embryonic development of the soldiers was studied by Ijichi et al. (2004).
P. bambucicola is found in Japan, Ryukyu, Taiwan, Hong Kong, Thailand, and northern Vietnam. South-east Asian populations are referable to P. carolinensis (Fukatsu et al. 2001). Aphids of the P. bambucicola/carolinensis group also occur in India, Sri Lanka and Fiji, but their specific identity has not been established. 2n=12.
Pseudoregma carolinensis (Takahashi) Galls on Styrax benzoides in northern Thailand are green, in clusters of up to ten petiolate sub-galls, formed from flower buds, similar to those of P. bambucicola but smaller, greener and flatter, and with a beak-like apical extension (Aoki et al. 2002) (see influentialpoints.com/Gallery). Gall populations include second instar soldier larvae. Alatae (BL 1.3-1.8 mm) maturing in early August fly to bamboos (Bambusa spp., Dendrocalamus giganteus). Apterae on bamboos are broadly oval, in life grey brown or dark purplish to greenish brown or black, dusted with granular wax: BL 1.5-2.6 mm. Colonies are usually formed on the culms or twigs. Individuals in young colonies may have three longitudinal rows (two marginal, one spinal) of white wax tufts, but in older colonies these are less developed and adults have a thick cushion of wax posterior to the siphunculi (Noordam 1991, as P. bambucicola). Adult apterae are apparently morphologically indistinguishable from those of P. bambucicola, but DNA fingerprinting has shown that P. carolinensis is a distinct species of more southerly distribution, occurring in Thailand, Malay Peninsula, Java, Irian Jaya and Micronesia (Fukatsu et al. 2001). These authors also reported a difference between first instars and soldiers of bambucicola and carolinensis in the number of hairs on abdominal tergite 8. First instar soldiers occur within the colonies on bamboo (Noordam 1991, as bambucicola), and alate sexuparae are produced in November-December, but parthenogenetic reproduction continues throughout the year, and secondary migrants (alatae dispersing between bamboos) occur in Java, and probably elsewhere (Aoki et al. 2002).
Pseudoregma dendrocalami (Takahashi) Green to brownish green apterae, with four wax dorsal longitudinal stripes, were described from Dendrocalamus flagellifer in Sumatra (Takahashi 1935a). Possibly P. albostriata Liao is this species.
Pseudoregma koshunensis (Takahashi) Galls on Styrax suberifolia in Taiwan are grey-brown, hard, banana- or flask-shaped, about 28 mm long with apical opening, occurring in bunches (Takahashi 1939; as Astegopteryx shitosanensis). Their inhabitants include many pseudoscorpion-like second instar soldiers (Aoki 1982). Inhabitants of the galls closely resemble the same forms of P. bambucicola and P. sundanica (Aoki et al. 2002). Heteroecious holocyclic; alatae migrate in June to Bambusa spp. (Aoki & Kurosu 1991b). Apterae infesting the culms or leaves of bamboo plants are broadly oval, purplish to greyish or dark brown, covered with white wax; BL 1.6-2.0 mm (Takahashi 1924a). Populations on various bamboos in mainland China have also been identified as this species (Qiao et al. 2018). Individuals on leaves are generally darker, more dorso-ventrally flattened, and the wax forms marginal tufts on posterior abdominal segments; they were described as a distinct species, pseudomontana (Takahashi 1924a), but most authors (e.g. Aoki & Kurosu 1991b) now regard the two forms as conspecific. Parthenogenetic populations may also persist through the year on bamboos.
Pseudoregma montana (van der Goot) Apterae are broadly oval, yellowish to violet-coloured with yellowish head and prothorax, dusted with wax and with four variably developed longitudinal rows of small white wax tufts (two spinal, two marginal); BL 1.9-2.8 mm (Noordam 1991). Immatures are bright yellow to orange, dusted with wax. In dense colonies on undersides of leaves near the leaf bases, of unidentified bamboos in Java. Large populations spread over the whole shoots. Apart from its size and colour, the difference between this species and P. koshunensis from Taiwan is unclear. [Indoregma bambusae Chakrabarti & Maity, described as apterae from Bambusa sp. in Uttar Pradesh, India (Chakrabarti & Maity 1982), is a Pseudoregma according to A.K. Ghosh (1988), who examined type specimens, and was synonymised by him with P. panicola, but that species is not a bamboo-feeder, and the proportions of the antennal segments and distribution of wax glands correspond much more closely to those of P. montana.]
Pseudoregma nicolaiae (Takahashi) Apterae are brownish black, secreting columns of white powdery wax in four longitudinal rows; BL 1.6-1.8 mm. On undersides of leaves, along veins towards middle of leaves (cf. sundanica), of various Zingiberaceae (Elettaria, Etlingera, Hedychium. In south-east Asia (Java, Malaya, Sumatra). It is closely related to P. sundanica (Stern et al. 1997). Defensive behaviour of soldiers was compared with those of P. sundanica by Shingleton & Foster (2001).
Pseudoregma panicola (Takahashi) Apterae are brownish black or dark brownish red-violet, secreting columns of dense white wax (see influentialpoints.com/Gallery); BL 1.1-1.9 mm. On many genera and species of tropical grasses (Andropogon, Arundo, Capillipedium, Cyrtococcum, Eragrostis, Ichnanthus, Indocalamus, Lasiacis, Oplismenus, Panicum, Paspalum, Pseudoechinolaena, Setaria, Stenthotheca, Thysanolaena), feeding on undersides of leaves, stems and inflorescences. In Africa, southern India, east and south-east Asia, New Zealand (Cottier 1953) and Australia (Brumley 2020), and also in Central America (Cuba, Venezuela, Costa Rica, Puerto Rico, Guadalupe; Villalobos Muller et al. 2010) (Mexico; Muñoz-Viveros et al. 2025). A record from Turkey on roots of Elytrigia (Görür et al. 2017) must be a misidentification. The life cycle and sexual morphs are unknown, and presumably it is mainly or entirely anholocyclic. Noordam (1991) provided full descriptions of known morphs.
Pseudoregma pendleburyi (Takahashi) Apterae are blackish brown, marbled; BL 1.3-2.7 mm. Originally described from Malaya on Bambusa sp. (Takahashi 1950), and redescribed (including first instar soldiers) from Java by Noordam (1991). Also recorded from West Irian (BMNH collection, leg. E.W. van Heurn). The populations in Java were living on the culms, but bore a close resemblance in size, colour and development of wax glands to the leaf-feeding population in Malaya that Takahashi (1950) described as a distinct species, gombakana. Development of wax glands in Pseudoregma depends greatly on age of colony and degree of alatiformity, and it seems probable that gombakana is a junior synonym of pendleburyi, which was described first in Takahashi’s (1950) paper. Oregma uscare Tao, described from unidentified bamboos in Okinawa (Tao 1964), is possibly also a synonym. The life cycle is unknown.
Pseudoregma sundanica (van der Goot) Galls on Styrax paralleloneura are up to 6.8 cm in diameter and consist of up to seven sub-galls arising from old twigs; velvety white with green internal projections visible through the slit-like openings (Kurosu & Aoki 2001). Numerous second instar soldiers occur within the galls. Alate emigrants from galls in early August were successfully transferred to found colonies on ginger (Etlingera elatior). The gall generations are only known from northern Sumatra, but secondary host generations are much more widely distributed in east and south-east Asia. Apterae on secondary hosts are blackish brown or black, sometimes with tufts of white wax marginally and spinally (see influentialpoints.com/Gallery); BL 1.1-2.0 mm. They occur on various Zingiberaceae (Alpinia, Amomum, Costus, Etlingera, Zingiber), colonising bases of leaves, stems and petioles, attended by ants (Noordam 1991), and are also recorded from Curculigo capitulata. In Java, Malaya, Sumatra, and Philippines, and introduced to Australia (Brumley 2020).
|
Pseudothoracaphis Raychaudhuri, Ghosh & Das |
Hormaphidinae: Nipponaphidini |
One species on Quercus in northern India and Pakistan, perhaps related to Allothoracaphis but lacking marginal prosomal hairs.
Pseudothoracaphis himachali Raychaudhuri, Ghosh & Das Apterae are oval, dark brown, dorso-ventrally flattened, strongly sclerotised; BL 1.1-1.6 mm. In ant-attended colonies on undersides of leaves of Quercus incana and Quercus sp. in India (Himachal Pradesh, Uttar Pradesh), and also recorded from Q. incana and Q. baloot in the north-eastern hills of Pakistan (Namann-Etienne & Remaudière 1995). The host plant of the holotype was reported to be Ilex sp., but this is likely to be an error. All specimens were collected in October (Raychaudhuri, L.K. Ghosh & Das 1980); other morphs and life cycle are unknown. Heminipponaphis querciphaga Chakrabarti & Raha (1985) seems to a synonym.
|
Pterasthenia Stroyan |
Pterastheniinae |
A small African genus mainly associated with woody Leguminosae. There are several records of the type species, Pterasthenia shiraensis Stroyan, from Poaceae, but these plants are almost certainly not the true hosts, and there are now records of this species from Millettia (see below). Remaudière & Quednau (1988) reviewed the genus, making Antalus a subgenus, and Quednau (2010) provided keys to species and illustrations of all available morphs.
Pterasthenia (Antalus) albata (Adams) Plate 6a Apterae have the head and prothorax reddish brown, rest of thorax and first two abdominal segments dark green, rest of abdomen yellow green, legs conspicuously wax‑powdered and body studded with clumps of white wax; BL 1.3-1.8 mm.. Alatae are brown and green with much reduced hindwings. On Indigofera spicata in central Africa (Burundi, Malawi, Zimbabwe, Tanzania). Common in the Central Province area of Malawi in the dry season, bcoming rare with the advent of the rains (Adams 1965).
Pterasthenia matileae Remaudière & Quednau All viviparae are alate, pale yellow with bluish white cottony wax on body, legs and antennae; BL 1.5-2.3 mm. Living all year on Milletia eetveldiana in the Congo Republic; colonising undersides of leaves in dry season, but in wet season found especially on young shoots at base of stem (Remaudière & Quednau 1988). Parthenogenetic reproduction continues throughout the year, and no sexual morphs are known. This aphid is attacked by a specific parasitoid (Trioxys pterastheniae; Stary et al. 1977).
Pterasthenia shiraensis Stroyan Alatae with woolly wax in life (J.H. Martin, pers. comm.) colour not recorded; BL 1.6-2.0 mm. Originally described from several samples from beneath grass tufts in Tanzania (Stroyan 1952), but there have subsequently been collections from Millettia sp(p). in Kenya and Tanzania (BMNH collection, leg. J.H. Martin, D. Hollis, Commonwealth Institute of Entomology), and from Congo, Uganda and Burundi (Quednau 2010). The biology is unknown.
|
Pterocallis Passerini |
Calaphidinae: Pterocallidini |
Thirteen species of small pale aphids with a short antennal terminal process living on Betulaceae, especially Alnus. They are usually dispersed on undersides of leaves, and are mostly not attended by ants. Apterous viviparae occur more commonly than in most Calaphidinae, particularly in Pterocallis s. str. which has four species in North America, three in Europe and three in east Asia. The subgenus Recticallis (three species) is wholly east Asian. Mesocallis is here considered as a separate genus, and we have followed Quednau (2003) in placing subgenus Paratinocallis under that genus. Accounts are available for Britain (Stroyan 1977), Fennoscandia and Denmark (Heie 1982), central Europe (Börner 1949), Japan (Higuchi 1972), India (Chakrabarti 1988, A.K. Ghosh & Quednau 1990), USA (Granovsky 1928a, as Myzocallis) and Canada (Richards 1965). Quednau (2003) provided keys and illustrations of all available morphs.
Pterocallis affinis Chakrabarti Alatae are pale yellow, with antennae and legs mainly pale; BL 1.5-2.0 mm. On Corylus colurna in Uttar Pradesh, India, and alatae have been trapped in Pakistan (Naumann-Etienne & Remaudière 1995). No apterae are recorded. Oviparae and males occur in October (Chakrabarti 1988).
Pterocallis albida Börner Apterae and alatae are yellowish white or greenish white, shining, with pale antennae and legs; BL 1.0-1.2 mm. On Alnus incana, dispersed on undersides of leaves. Widely distributed in continental Europe (see Lampel 1983 for details), and into Turkey (Görür et al. 2009). Oviparae and males occur in September-October (Italy; BMNH collection, leg. H.L.G. Stroyan, and Quednau 2003).
Pterocallis alni (De Geer) Apterae and alatae are yellowish white to yellowish green, with antennae ringed with black, a black spot near apex of hind femur, and black tarsi (see influentialpoints.com/Gallery); BL 1.3-2.0 mm. Almost exclusively on Alnus glutinosa in Europe, living dispersed on undersides of leaves and only occasionally attended by ants. Common and widespread in Europe, east to Turkey, Iran and Lebanon; introduced to North America (Essig 1912b), New Zealand (Lowe 1968), Australia (Carver 1980a) and Chile (Carrillo 1980). In North America it seems able to feed on various native Alnus spp. (BMNH collection). South-west Asian populations colonise A. orientalis and A, subcordata as well as A. glutinosa and tend to be intermediate in morphology between P. alni and P. maculata, so their specific identity is often uncertain. Oviparae and alate males occur in September-November. The male genitalia were described and illustrated by Wieczorek et al. (2011). Gange & Pryce (1990) investigated the effects of temperature and nutrition on development, and Gange & Llewellyn (1988) studied egg distribution and mortality. P. alni on alder windbreaks may be a useful alternative food for predators of orchard pests (Gange & Llewellyn 1989). 2n=20.
Pterocallis alnifoliae (Fitch) (fig. 28A) All adult viviparae are alatae and have a pale greenish yellow abdomen and darker yellow to orange head and thorax, antennae dark-banded, and distal halves of siphunculi black (see influentialpoints.com/Gallery); BL 1.3-1.7 mm. On undersides of leaves of Alnus rugosa and A. crispa in eastern North America. Oviparae and alate males occur in September-October. Earlier accounts of this species (e.g. Blackman & Eastop 1994) may include P. rhombifoliae, which has both apterous and alate viviparae.
Pterocallis (Recticallis) alnijaponicae (Matsumura) All adult viviparae are alate, yellowish with pale antennae and legs, abdomen with marginal rows of brown spots, and spinal tubercles sometimes also brown-tipped; BL 1.5-2.1 mm (Higuchi 1972). Recorded from various Alnus spp. in Japan, Korea, and China (Qiao et al. 2005b). Quednau (2003) illustrated an ovipara collected in Japan in late October.
Pterocallis essigi Quednau Colour in life is unknown; BL of apterae 0.8-1.1 mm, of alatae 1.0-1.4 mm. On Alnus rhombifolia in California (but also reported to occur on A. maritima in North Carolina, and on Alnus sp. in Maryland; Quednau 2003). Oviparae and apterous males occur in October-November. This includes the western material in Granovsky’s (1928) description of Myzocallis (now Pterocallis) rhombifoliae (see Quednau 2003).
Pterocallis heterophyllus Quednau Alatae are whitish to very pale yellow with blackish apices to antennal segments, a black spot at base of pterostigma, narrowly brown-bordered forewing veins and brown tips to dorsal body hairs; BL c. 1.6-1.7 mm. Apterae are unknown. Described from specimens collected on Corylus heterophylla in Korea in August and November (Quednau 1979). Sexual morphs and life cycle are unknown.
Pterocallis maculata (von Heyden) Apterae are yellowish green or green with a variably developed pattern of rather diffuse dark green dorsal cross-bands, black apices to antennal segments, a black spot near apex of hind femur, and black tarsi (see influential points/Gallery); BL 1.4-2.1 mm. Alatae have greenish abdomen with dark green markings. On undersides of leaves of Alnus spp., especially A. glutinosa, usually along main veins, almost always attended by ants (cf. alni). Widely distributed in Europe, east to Turkey and Iran, and now also recorded from Kazakhstan (Kadyrbekov 2018a). Apterous males and oviparae occur in September-October (Stroyan 1955, Quednau 2003).
Pterocallis montana (Higuchi) Alatae are pale yellow, with pale antennae, wings and legs; BL 0.8-1.2 mm. Apterous viviparae are unknown, and probably do not occur. On undersides of leaves of Corylus sieboldiana (Higuchi 1972; as Myzocallis montana) and Carpinus japonica (BMNH collection, leg. R.L. Blackman) in Japan, and also recorded from China (W. Zhang & Zhang 1994). Sexual morphs and life cycle are unknown. 2n=16 (Blackman 1986).
Pterocallis (Recticallis) nigrostriata (Shinji) (fig. 28B,C) All adult viviparae are alate, yellowish with black apices to antennal segments; a dark patch in middle of hind femur; forewings are characteristically pigmented with bands of fuscous bordering veins and lines of darker pigment between veins near wing-tip; black spinal tubercles on abdominal tergites 1-4, and black marginal spots on abdominal tergites 3-4; BL 1.4-2.0 mm. Immatures and oviparae have conspicuous long dark dorsal hairs. On undersides of leaves of various Alnus spp. in Japan (Takahashi 1965) and Korea. Paik (1972) recorded oviparae in Korea in early November. 2n=c. 26 (Blackman & Eastop 1994).
Pterocallis ostryae (Börner) Apterae and alatae are yellow; BL 0.9-1.4 mm (Börner 1949). Alatae have a dark spot at apex of hind femur and a dark triangle at base of Cu1b in forewing. On undersides of leaves of Ostrya carpinifolia in alpine and southern Europe (Austria, France, Italy), and Quednau (2003) also reports it from the Middle East. Quednau (2003) illustrated an oviparae collected in southern France in early September.
Pterocallis (Recticallis) pseudoalni (Takahashi) All adult viviparae are alate, with pale brown head and thorax and yellow abdomen; antennae banded with brown-black, forewing veins narrowly bordered with fuscous, dark hind legs, dark finger-like spinal tubercle on abdominal tergite 1, dark marginal spots on abdominal tergites 3-4, and black distal halves to siphunculi; BL 1.6-2.1 mm. On undersides of leaves of Alnus formosana in Taiwan, where it reproduces parthenogenetically throughout the year (Takahashi 1923; as Myzocallis pseudoalni). Takahashi (1924) provided detailed descriptions of all immature stages. Granovsky (1928) provided an illustrated redescription.
Pterocallis rhombifoliae (Granovsky) Apterae are pale green, pale yellow or whitish with darker green irregular patches (Granovsky 1928); BL 1.1-1.5 mm. Alatae are pale green or greenish yellow, the antennae ringed with black. On Alnus rugosa in eastern North America. Oviparae and alate males occur in October (Quednau 2003).
|
Pterochloroides Mordvilko |
Lachninae: Lachnini |
One species differing from other Lachninae in having a double row of large spinal tubercles.
Pterochloroides persicae (Cholodkovsky) Apterae are shiny, oval, dark brownish grey or grey with variably developed black dorsal markings and prominent black siphuncular cones (see influentialpoints.com/Gallery); BL 2.7-4.2 mm. Living along undersides of larger branches, and sometimes forming large colonies on trunks, of woody Rosaceae (especially Prunus spp.). Large amounts of honeydew are produced, and colonies are regularly attended by ants. In southern Europe, North Africa, south-west and central Asia, India, Pakistan and Indonesia; it appears to be gradually extending its range, with reports of economically damaging populations in Romania and Tunisia. Monoecious holocyclic on Prunus (persica, armeniaca, spinosa) in cooler regions according to Archangelsky (1917). Oviparae and an alate male (BL c.2.5 mm) collected in Iran at an altitude of 1590m were described and illustrated by Wieczorek et al. (2013a), and the male genitalia were described and illustrated by Wieczorek et al. (2012). Most populations seem to be anholocyclic, with evidence of two clonal haplotypes having different feeding positions on the host (Mdellel et al. 2012). Genetic variation in mitochondrial DNA (cyt b) sequence was studied in Tunisian populations by Kharrat et al. (2014). Morphometric variation among populations from five countries was studied by Mdellel & Kamel (2015). Ecological studies have been carried out in Egypt (Darwish et al. 1989), Italy (Ciampolini & Martorelli 1978), Lebanon (Talhouk 1977) and Yugoslavia (Velimirović 1976). Biological control has been attempted with Pauesia antennata (Rakhshani et al. 2005). 2n=20.
|
Pterocomma Buckton |
Aphidinae: Macrosiphini |
About 30 species of robust, hairy aphids living without host alternation on Salicaceae in the northern hemisphere. For many years treated as a separate tribe within Aphidinae, Pterocomma (and presumably the related genera Fullawaya, Paducia, Plocamaphis and Neopterocomma) have now been shown by molecular studies to be allied to Cavariella, Capitophorus and several other generaforming a separate clade within the Macrosiphini (Choi et al. 2017). They are evenly distributed between western palaearctic (9 spp.), eastern palaearctic (12 spp.) and nearctic (8 spp.). They tend to be sombre-coloured, but with often contrastingly brightly coloured siphunculi, and their colonies on the bark of branches and twigs are almost always attended by ants. Szelegiewicz (1965) and Wojciechowski (2003) reviewed the genus, and regional accounts are available for Britain (Stroyan 1984), Fennoscandia and Denmark (Heie 1986), Iberian peninsula (Nieto Nafría et al. 2002a), central Europe (Heinze 1961b), Indian region (Chakrabarti & Das 2008), east Siberia (Pashtshenko 1988a), China (Zhang & Zhong 1980), US Rocky Mountain region (Palmer 1952) and Canada plus Greenland (Richards 1967). Nowińska et al. (2017) provided a key to known males of European species. Tang et al. (2015) employed DNA barcodes for species identification, based on three molecular markers and comparing different methods of analysis. Specialised parasitoids include Aphidius pterocommae and perhaps also Aphelinus fulvus, which is also recorded from willow-feeding Chaitophorus.
Pterocomma alpinum Zhang, Zhong & Zhang Apterae are red-brown in life, with slight dusting of wax; BL c.3.4 mm. Alatae have 17-21 secondary rhinaria on ANT III. On an unidentified Populus sp. in Yunnan, China (G. Zhang et al. 1992c).
Pterocomma atuberculatum Chakrabarti & Das Colour of aptera in life is not recorded; BL 3.0-3.4 mm. Alatae have 15-16 secondary rhinaria on ANT III. On Populus nigra in Bhutan (Chakrabarti & Das 2008).
Pterocomma baicalense Ivanoskaya Apterae are dark grey-greenish with dark sclerites posterior to siphunculi; BL 2.9-4.0 mm. On Salix sp.(p.) in east Siberia (Baikal). Alatae have 9-19 rhinaria on ANT III. Wojciechowski (2003) provided a redescription. Contrary to Blackman & Eastop (2006), this species is clearly distinct from Pt. groenlandicum, having well-developed marginal tubercles, shorter R IV+V and longer siphunculi. It will run to couplet 28 in our key to Pterocomma spp. on Salix.
Pterocomma bailangense Zhang. Colour in life is not recorded; BL of aptera c. 4.6 mm. On an unidentified Populus sp. in Tibet (Zhang & Zhong 1981). Other morphs and biology are unknown.
Pterocomma beulahense (Cockerell) (fig. 84C) Apterae are yellowish olive-green (Palmer 1952), probably sometimes darker; BL 2.6-4.0 mm. Alatae have a grey-brown abdomen with black dorsal cross-bands. On twigs of Populus tremuloides in the Rocky Mountains, from Washington to New Mexico. Presumably holocyclic; a male was collected in September (BMNH collection, leg. D. Hille Ris Lambers). Richards (1967) synonymised beulahanse, which seems to be specific to P. tremuloides, with the more polyphagous Pt. bicolor, but the two species can be distinguished by the characters given in the poplar-aphid key.
Pterocomma bhutanense Chakrabarti & Das Appearance in life is unrecorded; BL of aptera 3.1-3.6 mm. Alatae have 20-26 secondary rhinaria on ANT III. On Populus nigra in Bhutan (Chakrabarti & Das 2008).
Pterocomma bicolor (Oestlund) (fig. 84B, fig. 106O) Apterae are reddish brown to dusky greenish yellow, greyish or bluish green, with yellow-orange siphunculi (see influentialpoints.com/Gallery); BL 3.2-4.8 mm. Alatae have a reddish brown to dusky green abdomen with variably-developed dark brown to black dorsal markings, only sometimes forming complete cross-bands. On bark of twigs and small branches, or on trunks of saplings, where they occasionally feed just below ground level. Native host plants are Populus spp. (especially angustifolia, balsamifera, tremuloides, trichocarpa), but this species also occurs commonly throughout North America on naturalized Salix spp. (alba, babylonica). Widespread in North America, and in Mexico. Oviparae and alate males occur in October-November. 2n=8.
Pterocomma (Stauroceras) chaetosiphon (Börner) (fig. 105E, fig. 106I) Apterae are greenish grey to brownish, with yellow siphunculi; BL 2.5-3.0 mm. Alatae have not been described. In bark crevices of trunk and branches of Salix spp. (fragilis, alba) in central Germany (Heinze 1961), and also recorded from Ukraine (Zhuravlev 2001). Oviparae have been collected in late September (BMNH collection, leg. D. Hille Ris Lambers).
Pterocomma dubium Börner Apterae are silvery-grey, dusted with wax; BL 2.6-3.2 mm. Fundatrices have BL c. 4.3 mm. In colonies on young shoots of Populus alba, never attended by ants (cf. populeum). There are also records from P.× generosa (Cambridge, England, BMNH collection), and P. laurifolia (Mongolia, Holman & Szelegiewicz 1974a, as P. populeum dubium). Probably widely distributed in Europe, and in Mongolia. Oviparae and alate males occur in late September-October; Wojciechowski (2003) described or redescribed all morphs. [This aphid has been synonymised in catalogues with Pt. populeum, but apart from the absence of marginal tubercles there are other consistent differences (colour in life, feeding site, host preference and lack of ant attendance) indicating separate species status.]
Pterocomma groenlandicum Hille Ris Lambers Colour in life is unknown; BL of aptera 2.8-3.7 mm. Alatae have well-developed dark dorsal abdominal cross-bands. On Salix sp. in Greenland, and also now recorded from Wrangel Island, East Siberia by Stekolshchikov & Khruleva (2014), who described a putative fundatrix, collected from S. lanata in early July. A record from Turkey (Şenol et al. 2017) requires additional confirmation. Oviparae and alate males occur in Greenland in late July-August (Hille Ris Lambers 1952). Khruleva & Stekolshchikov (2019) redescribed the ovipara (from East Siberia). Closely related to Pt. bicolor, of which it was placed as a subspecies by Richards (1967).
Pterocomma italica Barbagallo & Stroyan (fig. 106L) Apterae are very dark brown, with a fine spinal line of whitish wax crossed by two transverse wax bands on about abdominal tergites 1 and 5, and orange-yellow siphunculi;, BL 2.3-3.4 mm. Alatae have variable dorsal abdominal markings, not forming cross-bands. On twigs or at base of stems of Salix purpurea, in mountainous areas of Italy and Switzerland (Barbagallo & Stroyan 1978). Sexual morphs and life cycle are unknown. Very similar to Pt. jacksoni. Heavy parasitisation by an Aphidius species is reported in the original description.
Pterocomma jacksoni Theobald (fig. 106K) Apterae vary in colour from pinkish or reddish brown to brown-black, with a greenish tinge, sometimes have fine lines of wax as in italica and salicis, and orange-yellow siphunculi; BL 2.2-4.3 mm. Alatae are without dorsal abdominal cross-bands. On branches or stems of Salix spp. (argyracea, aurita, caprea, cinerea, repens), forming dense ant-attended colonies either above or below ground level. Widely distributed in Europe (see Heie 1986), but uncommon; also in western Siberia and Kazakhstan (Kadyrbekov 2017a). Sexual morphs and life cycle are unknown. 2n=c.30-34 (Kuznetsova & Shaposhnikov 1973).
Pterocomma konoi Hori Apterae are greyish or brownish, with yellow siphunculi; BL 3.0-4.0 mm. In dense ant-attended colonies along stems and branches of many Salix spp. Alatae have 22-38 secondary rhinaria on ANT III (cf. pilosum), and 0-3 on ANT IV. Widely distributed through Europe and Asia. Oviparae, occurring on S. caprea in September-October (in Sweden) have darker and more swollen hind tibiae with many more scent glands than Pt. pilosum, of which it is often placed as a subspecies (Heie 1986). In Europe, konoi may occur in mixed colonies with pilosum, indicating that there may be an effective isolating mechanism, and that the two taxa are probably distinct species.
Pterocomma kozchukovae Pashtshenko Apterae are orange-brown, brown or brownish-green, dusted with wax; BL c. 3.5 mm. Forming dense ant-attended colonies on stems and suckers of Populus bolleana (= P. alba var. pyramidalis) in maritime east Siberia. Holocyclic, with alate males (Pashtshenko 1988a).
Pterocomma lhasapopuleum Zhang Colour in life is not recorded; BL of aptera c. 3.7 mm. Alatae have dark dorsal abdominal cross-bands. On an unidentified Populus sp. in Tibet (Zhang & Zhong 1981b). Sexual morphs and life cycle are unknown.
Pterocomma pilosum Buckton (fig. 106N) Apterae are greenish, greyish or brownish, with yellow siphunculi (see influentialpoints.com/Gallery); BL 2.3-4.0 mm. Alatae have dark dorsal abdominal cross-bands, sometimes divided medially into paired patches, and secondary rhinaria distributed ANT III 33-44, IV 0-5 (cf. konoi). On twigs and branches or in bark crevices of numerous Salix spp., usually attended by ants. In Europe and Asia (western Siberia, Iran, Kazakhstan, Kadyrbekov 2011c); Kyrgyzstan, Kadyrebekov 2013c; Pakistan, Naumann-Etienne & Remaudière 1995; Inner Mongolia, Wang et al. 2013). There is also a record from Algeria (Laamari et al. 2013). Introduced to western North America (Utah, British Columbia). Oviparae and alate males occur in October. Szelegiewicz (1967a) described a variant population, from Salix alba var. pendula in Poland, as a subspecies Pt. pilosum sarmaticum; similar specimens were also found on S. alba in Netherlands (BMNH collection, leg. H.L.G. Stroyan). [Moritsu’s (1983) account and illustration of an aphid with long fine hairs on the siphunculi under the name pilosum must refer to some other, undescribed, Japanese species.] 2n=8.
Pterocomma populeum (Kaltenbach) Plate 12e, f Apterae are yellowish grey or brownish, with paired dark patches at least on posterior abdominal tergites, intersegmental cross-bands of greyish white wax, and pale siphunculi (see influentialpoints.com/Gallery); BL 2.7-4.5 mm. Alatae have broad dark dorsal abdominal cross-bands. Ant-attended, on branches or two-year-old twigs, or in bark crevices, of many Populus spp., but especially common on P. nigra and its varieties. Widespread in Europe, including Spain, and also recorded from North Africa, south-west and central Asia, India, Bhutan and east Siberia. Probably also in China, as Pt. kormion Zhang, Chen, Zhong & Li (in G. Zhang 1999) is likely to be a synonym. Introduced to North America (see Smith & Parron 1978, Forbes & Chan 1986a), South America (Argentina: Molinari 1942, and Chile: BMNH collection, leg. E. Zuniga), and Australia (Brumley et al. 2024). Oviparae and alate males occur in October-November. The male genitalia were described and illustrated by Wieczorek et al. (2011). Martín-Bernal & Padro Simarro (1986) reviewed natural enemies. [There are numerous records of this species from Salix, but these are almost certainly due to confusion with other species (pilosum and konoi in the palaearctic; ?sanguiceps in North America.]
Pterocomma populifoliae (Fitch) Apterae are pinkish to brown, variably dusted with greyish wax, with siphunculi pale to dusky orange (Richards 1967) (see influentialpoints.com/Gallery); BL 2.2-3.8 mm. Alatae have variably developed dorsal dark patches, not forming cross-bars except on posterior tergites. On Populus spp. (balsamifera, grandidentata, tremuloides) in north-eastern USA and Canada, usually colonising roots or at bases of stems of sapling trees. Oviparae and alate males occur in September. This species is generally confused in the North American literature with the widely distributed and more polyphagous Pt. bicolor (Richards 1967); Palmer’s (1952) account of populifoliae should be referred to that species. Occurrence of Pt. populifoliae in India is based on sexual morphs collected on Populus ciliata (Saha & Chakrabarti 1988, Chakrabarti & Das 2008) and needs further confirmation.
Pterocomma pseudopopuleum Palmer Apterae are greyish to yellowish olive-brown, finely dusted with wax; BL 3-4 mm. Alatae have broad dark dorsal abdominal cross-bands. On bark of twigs and small branches of Populus spp. (especially angustifolia and deltoides ssp. occidentalis), in the Rocky Mountain region (Idaho, Utah, Colorado, New Mexico). Oviparae and alate males occur in October-November (Palmer 1952).
Pterocomma rufipes (Hartig) (= steinheili Mordvilko) (fig. 106P) Apterae are variably pigmented, grey or dull reddish brown to dark brown with spots of powdery wax and yellowish siphunculi (see influentialpoints.com/Gallery); BL 2.9-4.6 mm. Alatae have variably developed dorsal abdominal cross-bands. On twigs and young branches of numerous Salix spp, attended by ants. Heie (1986) recorded it also on ornamental and hybrid balsam poplars. In north-west, north and central Europe, Iceland, west Siberia, Turkey (Akyürek et al. 2019), Uzbekistan (Kadyrbekov 2013c), Kazakhstan (Kadyrbekov 2017a), Mongolia, India, east and north-east Siberia (Pashtshenko 1988a, Stekolshchikov & Khruleva 2015), and also in Canada (Manitoba; BMNH collection, leg. A.G. Robinson). Oviparae and alate males occur in September. Larger, darker specimens with longer siphunculi are sometimes regarded as a separate species, Pt. ringdahli, but are most probably part of a range of intraspecific variation (Heie 1986). Pt. henanense Zhang, on Salix sp. in China (Zhang & Zhong 1980), and Pt. tibetasalicis Zhang, on Salix sp. in Tibet (Zhang & Zhong 1981), are closely related, if not synonyms. 2n=8 or 9 (Kuznetsova 1974, as steinheili).
Pterocomma salicicola (Uhler) The identity of this species is uncertain. It could possibly be an earlier name for P. smithiae.
Pterocomma saliciphagum Chakrabarti & Das Appearance in life is unrecorded; BL of aptera 2.7-3.2 mm. Apterae and immatures were collected from an unidentified Salix sp. in Bhutan (Chakrabarti & Das 2008). Alatae and other morphs are unknown. Very similar to Pt. konoi; it needs to be confirmed that the differences are not due to size or seasonal variation.
Pterocomma salicis (Linnaeus) (fig. 106J) Apterae and alatae are greenish black to black, marked dorsally with greyish white wax powder including a spinal stripe and lateral spots, and bright orange siphunculi (see influentialpoints.com/Gallery); BL 2.7-4.5mm. Forming dense colonies on two-year-old twigs and wands of Salix spp, attended by ants. Widely distributed in Europe and Asia; introduced and widespread in North America. Apterous males and oviparae occur in October-November. Holman & Szelegiewicz (1974a) discriminated a subspecies, Pt. salicis rohdendorfi, on Salix sp. in Transbaikalia and Mongolia. 2n=58 (Blackman & Eastop 1994).
Pterocomma salijaponica (Shinji) Apterae are orange or brown with orange siphunculi (Shinji 1924, 1927); BL 2.8-3.5 mm. Alatae have dark dorsal abdominal cross-bands, sometimes broken in mid-line. On stems of Salix spp (babylonica, gracilistyla, koriyanagi) in Japan. Monoecious holocyclic, with alate males (Shinji 1941). The identity of this species is uncertain; it could not be included in the key to Salix-feeders on the basis of the available information. It was listed in error as a Plocamaphis by Eastop & Hille Ris Lambers (1976), and Paik’s (1965) account under Clavigerus salijaponica is of Plocamaphis coreana. The karyotype is also uncertain; 2n=8 or 2n=22 (Shinji 1927, 1931, 1942 – see Blackman 1986, as Plocamaphis salijaponica).
Pterocomma sanguiceps Richards (fig. 106M) Apterae have head and prothorax bright red, rest of dorsum bluish, with yellow to pink siphunculi; BL 3.0-3.8 mm. Alatae have abdomen brownish or bluish, without dark dorsal cross-bands. On Salix spp (babylonica, exigua) in British Columbia, Canada (Richards 1967; BMNH collection, leg. C. K. Chan). 2n=8 (Blackman & Eastop 1994).
Pterocomma sanpunum Zhang Apterae are yellowish red; BL c. 3.9 mm. On Populus cathayana and P. simonii in China (Zhang & Zhong 1980b), and also recorded from P. laurifolia and P. talassica in Kazakhstan (Kadyrbekov 2009a, 2014d). Other morphs and life cycle are unknown. Closely related to, if not synonymous with, Pt. yezoense.
Pterocomma smithiae (Monell) (fig. 84A) Apterae are dark reddish brown with bluish pruinosity and yellow siphunculi (see influentialpoints.com/Gallery); BL 2.3-4.0 mm. Alatae are without dark dorsal abdominal cross-bands. On bark of twigs of numerous Populus and Salix spp, common and widely distributed through North America, and in Mexico. Oviparae and alate males occur in October-November (Palmer 1952, Richards 1967).
Pterocomma tremulae Börner Apterae are dark warm brown or olive-brown, wax-dusted, with straw-coloured siphunculi (see influentialpoints.com/Gallery); BL 2.5-4.3 mm. Alatae have variably developed dark dorsal abdominal sclerites, usually divided into paired patches and not forming complete cross-bands, and secondary rhinaria distributed III 45-54, IV 0-7. Usually on suckers and two-year-old twigs of Populus tremula, but occasionally found on other Populus spp (see Heie 1986). Attended by ants. In north-west, northern, central and eastern Europe, west Siberia (Stekolshchikov et al. 2008a) and Kazakhstan (Kadyrbekov 2017a). Oviparae and alate males collected in October were described by Nowińska et al. (2017). 2n=8.
Pterocomma xerophilae Ivanovskaya Apterae are greenish yellow to brownish; BL c. 2.1 mm. On twigs of Salix xerophila and Salix sp. in west Siberia, on Salix sp. in Kyrgyzstan (Kadyrbekov 2013c), and on various Salix spp. in Kazakhstan (Kadyrbekov 2005c). Apterae have well-developed wart-like marginal tubercles (Ivanovskaya 1977). Monoecious holocyclic; an alate male was recorded in late September (Ostanin 1976).
Pterocomma yezoense (Hori) Apterae are orange-red to reddish brown, with antennae and legs mostly dark, and siphunculi pale yellow with dark apices; BL 2.9-3.8 mm. Alatae have dark dorsal abdominal cross-bands, sometimes broken in the mid-line. On suckers, young stems and branches of Populus spp. in Japan, maritime east Siberia (Pashtshenko 1988a) and China (BMNH collection, leg V.F. Eastop). Oviparae and alate males occur in Noember (Hori 1929b). Several other Pterocomma species described from Populus in China (anyangense, lazikouense, monticulum, neimongolense, sinipopulifoliae) are very similar (G. Zhang & Zhong 1980b; G. Zhang 1999), and their separate identity needs further confirmation. 2n=8 (Blackman & Eastop 1994).
|
Pyrolachnus Basu & Hille Ris Lambers |
Lachninae: Tuberolachnini |
A small genus of very large east Asian aphids living on woody Rosaceae. The alatae have an elongate pterostigma as in Longistigma, but it does not curve around the tip of the wing. A.K. Ghosh (1982b) reviewed the species in India.
Pyrolachnus imbricatus David, Narayan & Rajasingh Apterae are dull brown with transverse rows of dark brown spots on dorsum, and blackish tibiae and tarsi; BL 4.4-5.2 mm (David et al. 1971d). Alatae have only 9-15 secondary rhinaria on ANT III (A.K. Ghosh 1984). Feeding on stem and branches of Prunus spp., including P. cornuta (Medda et al. 1986). In India (Himachal Pradesh, Uttar Pradesh). Sorin (2011) described a population on twigs of P. yedoensis in Japan as a subspecies, P. imbricatus nipponicus. The apterae of this form are smaller (BL 2.7-3.0 mm) and have other significant differences that led Kanturski & Lee (2024) to move it to a new genus as Miyalachnus nipponicus.
Pyrolachnus pyri (Buckton) Apterae are dull yellow to dark brown, often dusted with wax meal, antennae and legs blackish; BL 3.5-6.0 mm. Alatae have wings dark at their bases, and secondary rhinaria distributed III 11-53, IV 2-11, V 0-3 (variation perhaps reflecting degree of alatiformity). On bark of branches of Pyrus communis, and also recorded from Malus domestica and Eriobotrya japonica. In Turkey, Iran, Bahrain, Pakistan, India, Nepal, Sri Lanka, Korea and China. Holocyclic in China, where regular spring and autumn migrations suggesting host alternation were observed (Long & Chen 1988). Apparently anholocyclic elsewhere. P. macroconus Zhang, described from Eriobotrya in China (Zhang & Zhong 1982d) is closely related and possibly a synonym.